
doi: 10.1007/s11259-025-10736-6.
- PMID: 40227349
摘要
本研究的目的是调查巴西城市蝙蝠中重要人畜共患病原体的存在,并评估被诊断出狂犬病的蝙蝠中是否存在共同感染。2021年,使用来自巴西东南部城市地区52只蝙蝠的器官样本,诊断该地区的狂犬病和其他人畜共患病。阳性样本进行测序、表征,并录入GenBank。在圣曼努埃尔市一户人家收集到的一只黑色蝙蝠中发现了涉及狂犬病病毒、贝氏巴尔通体和利什曼原虫的共同感染,该市是内脏利什曼病的疫区。系统发育分析显示获得的RABV序列与人类和野生犬科动物的参考序列相似,并确认了检测到的巴尔通体和利什曼物种的身份。据我们所知,这是首次报告一只蝙蝠同时感染狂犬病病毒、贝氏巴尔通体和利什曼原虫。三种重要病原体在黑色蝙蝠中的共同感染强调了新热带蝙蝠作为人畜共患病原体宿主的多元作用。这一前所未有的发现加强了这些动物在病毒、细菌和原虫病原体传播动态中的潜在作用,以及在蝙蝠中进行分子监测的重要性。
关键词 RABV·巴通氏菌病·利什曼病·翼手目·宿主·分子监测
引言
人畜共患病对全球公共卫生构成重大挑战,特别是在生物多样性丰富和热带地区(Allen等人,2017年)。人类与野生动物之间日益增长的互动,受城市化、森林砍伐和环境失衡的驱动,是导致人畜共患病出现和传播的主要因素之一(Jones等人,2008年)。蝙蝠因其广泛的遗传和生态多样性,作为各种病原体的宿主发挥着至关重要的作用,包括病毒、细菌和原虫(Alexander等人,2018年;Franca等人,2024年)。
狂犬病、巴通氏菌病和内脏利什曼病是在巴西发生的人畜共患病。狂犬病在某些地区是地方性的,尤其是在农村和森林地区,目前已被认为是主要的感染源,将病毒传播给人类和其他动物(Mayen 2003年)。根据巴西卫生部的数据,从2010年到2024年,巴西报告了大约48例人狂犬病病例。其中,24例是由蝙蝠引起的(占所有病例的一半),9例由狗咬伤引起,6例由非人灵长类动物引起,2例由狐狸引起,4例由猫引起,1例由牛引起。在2例中,无法确定引发攻击的动物(Brasil 2024年)。巴托洛米病是一种在该国诊断不足的动物园诊断疾病,主要与宠物身上的跳蚤有关,在最近的流行病学研究中显示出高发病率(池田等人,2017年;冈瓦雷斯-奥利维拉等人,2020年;雷蒙多等人,2022年;法奇尼-马丁内斯等人,2023年;弗兰卡等人,2025a年)。在2011年至2017年间,37.94%(480/1265)的疑似病例得到确诊,城市人口比农村人口受影响更严重(席尔瓦等人,2019年)。内脏利什曼病在该国的几个地区是地方性的,巴西占全球病例的93%(德朱斯,2004年)。每年发生超过50万例(德朱斯,2004年)。它通过经常与人类、家畜和野生动物互动的白蛉传播(戈麦斯-埃尔南德斯等人,2017年;卡斯特罗等人,2020年;里瓦等人,2021年;弗兰卡等人,2025b年)。
蝙蝠被认为是众多人畜共患病原体的重要宿主(韩等人,2015年;古普塔等人,2021年)。尽管蝙蝠被认为是多种病原体的宿主,但关于这些动物中共感染的发生情况了解甚少,特别是与人畜共患疾病重要性有关的病原体。了解蝙蝠中人畜共患病原体的共同循环可能为溢出事件的潜在风险提供洞见(布鲁克和多布森,2015年)。本研究的目的是调查巴西城市蝙蝠中重要的人畜共患病原体的存在,并评估被诊断出狂犬病的蝙蝠中是否存在共感染。
材料与方法
2021年,从城市地区提交了52只蝙蝠样本进行狂犬病监测,随后调查了该地区发生的其他被忽视的人畜共患病原体(巴尔通菌属和利什曼原虫)。根据巴西卫生部的建议,对蝙蝠进行狂犬病的被动监测是国家环境和流行病学监测服务的关键组成部分,由地方卫生当局负责执行(巴西2022年)。表现出异常行为的蝙蝠,如攻击性或在白天飞行,或者在家中被发现的,会被市动物传染病服务部门收集并进行狂犬病诊断。当居民在家中发现蝙蝠时,被告知要联系这些部门。蝙蝠种类的鉴定基于形态特征,如体型大小、翅膀形状和颅骨结构,按照Reis等人(2013年)描述的标准进行。收集到的蝙蝠种类有:大菊头蝠(18只)、灰鼻蝠(15只)、北美蝙蝠(9只)、大棕蝠(3只)、南蝠(2只)、大菊头蝠(1只)、红蝙蝠(2只)、巴西蝙蝠(1只)和布氏黄鼠狼(1只)。
动物通过在封闭室内暴露于5%异氟醚浓度下安乐死,随后持续暴露直至心肺骤停。之后,在无菌环境下对动物进行尸检。最初打开颅骨取出大脑,然后取出脾脏和肝脏。在尸检过程中,对蝙蝠进行了彻底检查,以识别可能存在的异常或与动物源病原体感染相关的迹象。检查重点关注宏观病理学迹象,这些迹象表明可能患有动物源感染,包括器官变色、坏死、肉芽肿或非典型液体积聚。所有受检组织均在正常参数范围内。脑组织完全用于狂犬病的即时诊断,而其他组织则储存在无菌试管中,并在-80°C下冷冻保存,以供后续分析其他动物源病原体。
为了检测狂犬病毒,使用脑组织碎片进行直接荧光抗体试验(DFAT)和鼠接种试验(MIT)。DFAT涉及将荧光标记的抗体应用于脑组织印片上,通过荧光显微镜直接检测脑组织中的RABV抗原(Rupprecht等人,2018年)。MIT涉及将捣碎的脑组织注入健康小鼠体内,并监测它们七天内的临床症状,以确认RABV的存在(Rupprecht等人,2018年)。使用制造商提供的方案,按照InviSorb® Spin Universal Kit(Invitek Molecular公司,柏林,德国)说明书,从脑组织碎片中提取病毒RNA。提取的RNA作为通过逆转录合成cDNA的模板,使用的引物为21 g(5′-ATG TAA CAC CTC TAC AAT G-3’)(Menozzi等人,2020年)。如在其他地方所述(Menozzi等人,2020年),使用引物21g和304(5′-TTG ACG AAG ATC TTG CTC AT- 3’)扩增cDNA,目标是N基因的1,478个碱基对的片段。
为了检测巴通体,使用与RNA提取相同的试剂盒从脾脏样本中提取DNA。实时PCR作为筛选方法进行,目标是编码柠檬酸合成酶的338个碱基对的基因片段(gltA),如在其他地方所述(Buss等人,2012年)。使用的正向引物为BartGltAF(5′-TGC AGG WTC ATC RGG WGC TAA T- 3’),反向引物为BartGltAR(5′-TGA GCT GGT CCC CAA AGG- 3’)和BartGltAR2(5′-TGG CTC CAC CAT GTG CGG-3’,按1:1比例混合)。为了增强反应并提高对巴通体物种的特异性,使用了Bartpr探针(5′-FAM-CAT TTG CAT GTA TTG CAG CAG GTG TTG C-BQHI-3’)。作为阳性对照,使用了由米纳斯吉拉斯州公共卫生实验室的立克次体病和汉坦病毒服务提供的汉赛巴通体分离株的DNA。确认阳性后,使用引物BhCS.781p(5′-GGG GAC CAG CTC ATG GTG G- 3’)和BhCS.1137n(5′-AAT GCA AAAAGA ACA GTA AAC A- 3’)通过常规PCR扩增样本,目标是编码750个碱基对柠檬酸合成酶的基因,如在其他地方所述(Norman等人,1995年)。
为了检测利什曼原虫,使用了从脾脏样本中提取的DNA。实时PCR作为筛选方法进行,目标是潜在的腐胺合成酶1基因的204个碱基对的片段,如在其他地方所述(Wu等人,2022年)。正向引物CL-SPDF(5′-AGA TCA TTG CGT ACT TGA C- 3’)和反向引物CL-SPDR(5′-TCA TCG ACA CAA CAG ACC- 3’)被使用。使用了由圣保罗州立大学寄生虫学实验室提供的利什曼原虫MHOM/BR/IOC/L2906菌株的DNA作为阳性对照。确认阳性后,使用引物Leish_18S/ITS1(5′ CTG GAT CAT TTT CCG ATG ATT A 3’)和Leish_18S/ITS1(5’AAC ACA AAC ACA CGC CTC CT 3’)通过常规PCR扩增样本,这两个引物针对的是18S基因的530个碱基对片段以及ITS1核糖体RNA基因的开始部分,如参考方法(Franca等人,2025b年)所述。
然后将10微升PCR产物进行琼脂糖凝胶电泳。
阳性PCR产物使用Wizard®基因组DNA纯化试剂盒(Promega公司,美国威斯康星州麦迪逊市)按照制造商指南进行纯化,并送往进行桑格测序。阳性样本的序列与GenBank数据库中先前提交的序列使用BLASTn工具(Altschul等人,1990年)进行比较。序列质量使用BioEdit序列比对编辑器第7.2版(Ibis Therapeutics公司,美国加州卡尔斯巴德市)进行检查。
系统发育分析使用MEGA12版本12.0.7(Tamura等人,2021年)中的最大似然法进行。应用了带伽马分布的Tamura三参数模型(T92+G),并通过1000次自助法复制评估推断出的树拓扑结构(Tamura 1993年)。从狂犬病病毒分离株获得的序列与一个包含20个巴西N基因序列的数据集进行了比对,这些序列代表不同的宿主,包括狗、猫、马、牛、野生犬科动物、人类、非人灵长类动物和不同种类的蝙蝠。从巴尔通体属和利什曼原虫获得的序列与全球参考序列进行了比较。选择了外群来作为树的根:来自欧洲蝙蝠的狂犬病病毒、用于巴尔通体分析的不孕棒状杆菌以及用于利什曼原虫分析的锥虫。
结果
在分析的蝙蝠中,1.9%(1/52)对狂犬病病毒呈阳性,3.9%(2/52)对巴尔通体属细菌呈阳性,5.8%(3/52)对利什曼原虫呈阳性。在一只黑鼠耳蝠个体中观察到狂犬病病毒、巴尔通体和利什曼原虫的共同感染。这只蝙蝠是一只成年雌性。
实时PCR检测到的阳性样本的循环阈值(Ct)范围在30到32之间,表明微生物负荷适中。阳性样本的熔解温度(Tm)值与阳性对照一致,最大变化为1°C。
所有样本都通过常规PCR成功扩增,产生了足够的DNA进行测序。测序样本的查询覆盖率超过80%,且与其他GenBank中可用的序列相似性超过97%。共同感染的黑鼠耳蝠的序列登录号分别为PQ812415、PQ812416和PP939070,分别显示出与狂犬病病毒、科勒巴尔通体和小利什曼原虫的高度相似性。
如图1所示的系统发育分析揭示,从蝙蝠黑面鼯鼠(PQ812415)获得的狂犬病毒(RABV)的N基因序列与人类(Homo sapiens)和野生犬科动物(Lycalopex vetulus)的进化枝关系较近。图2展示了该基因的遗传信息。
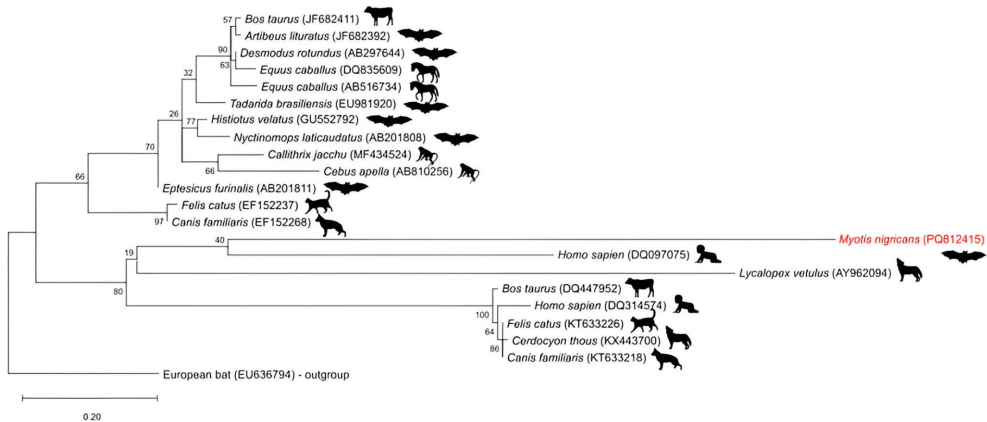
使用三参数模型和1000个自助法复制样本以评估分支支持度。使用来自欧洲蝙蝠的狂犬病病毒序列作为外群。
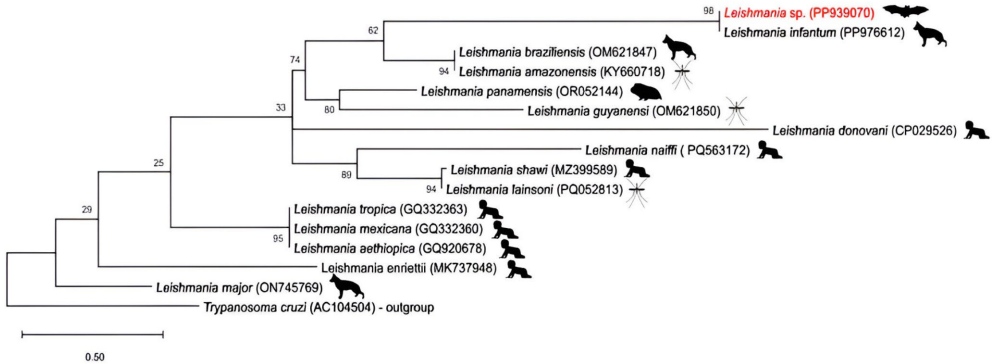
我们的序列(PQ812416)与B. koehlerae参考序列(MT095046)之间的相似性,确认了检测到的巴氏杆菌的身份。图3显示了我们的序列(PP939070)与利什曼病参考序列(PP976612)之间的遗传相似性,确认了检测到的利什曼原虫的身份。
讨论
在一群蝙蝠中检测到的狂犬病病毒、B. koehlerae和利什曼原虫的共感染代表了一个前所未有的发现。这一结果强调了这些动物同时携带多种具有重大公共卫生意义的人畜共患病原体的能力。本研究中阳性的蝙蝠属于圣马利亚市(西经48°27’4.91英寸,南纬22°55’41.45英寸),位于巴西南部圣保罗州内陆。该地区被巴西大西洋森林覆盖,这是一个重要的生态热点,以生物多样性高和城市化的强烈共存为特征(奥利维拉 2007年)。这种蝙蝠通常被称为黑叶鼻蝠,是一种广泛分布于拉丁美洲的食虫蝙蝠,能在从森林区域到城市环境的各种栖息地中找到,以其小巧的体型和深色的毛皮为特征(威尔逊 1971年)。
2017年,对一个包含超过2800种哺乳动物病毒感染的数据库进行了分析,发现蝙蝠是此类感染受影响最严重的物种(奥利瓦尔等人 2017年)。据报道,狂犬病毒已在41种巴西蝙蝠中发现,它们现在被认为是城市地区狂犬病发生的主要传播媒介(Menozzi等人,2020年)。黑蝙蝠能够长途迁徙寻找食物,并栖息于各种庇护所,通常与人类和家畜近距离接触。这种行为增加了接触和传播人畜共患病原体的可能性(Wilson 1971年;Ferreira等人,2018年)。这种动物不仅被狂犬病毒(RABV)感染,还被其他病原体感染,这突显了其存活感染的能力,并强调其作为其他重大疾病潜在宿主的角色。鉴于黑蝙蝠常在人类活动区域栖息,这种共同感染引发了对重叠传播途径(例如,媒介传播与直接接触)的担忧,并强调了在人类、家畜和蝙蝠交汇的分散栖息地需要进行综合监测的必要性(Ferreira等人,2018年)。
巴西大西洋森林是一个热带雨林区域,特点是高湿度和高温,这些条件不仅有利于丰富多样的动物物种生存,还有利于吸血蠓的繁殖(Pinto等人,2010年)。这些昆虫在自然和人造庇护所中与蝙蝠互动,以它们的血液为食,并充当病原体传播的媒介(Lampo等人,2000年)。在巴西,这些昆虫传播利什曼原虫属(Leishmania)的物种。
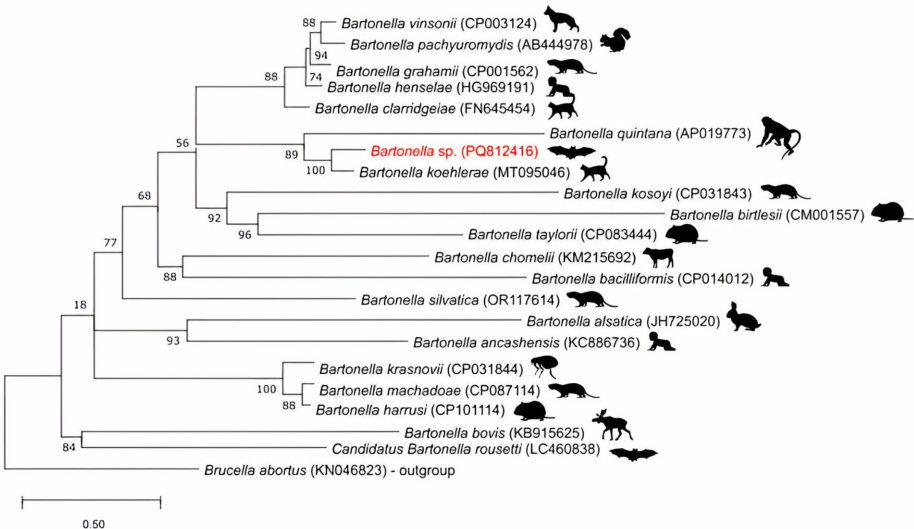
在MEGA12软件中使用似然法,采用Tamura 3参数模型和1000次自助法复制来评估分支支持。使用布氏杆菌作为外群。
最近的研究表明,巴特氏菌属(Bartonella)物种也能通过它们传播(李等人,2024年)。此外,美国的一项研究评估了食虫蝙蝠对库蚊的捕食情况,这暗示了向蝙蝠传播的另一种可能途径(休斯等人,2022年)。这些发现表明,白蛉可能有助于蝙蝠的共同感染。
收集蝙蝠的城市是内脏利什曼病的疫源地。此前已在巴西疫源地的蝙蝠体内检测到利什曼菌属物种;然而,尚未报告与其他病原体的共同感染(拉茨劳夫等人,2023年)。尽管该市尚未报告巴尔通体病病例,但该疾病在巴西仍被严重低估,尽管已有记录显示其发生。东南部最近的研究表明,在庇护猫、无家可归者和献血者中巴尔通体感染率很高(皮塔西等人,2015年;雷蒙多等人,2022年;法奇尼-马丁内斯等人,2023年)。家猫作为潜在的传播者,通常在受到跳蚤媒介感染后通过抓挠传播细菌。尽管猫的临床症状通常较轻,但在人类中,该感染可能导致严重疾病,如心内膜炎(阿维多尔等人,2004年)。此前已在巴西新热带蝙蝠体内报告过巴特氏菌的存在;然而,尚未报告与其他病原体的共同感染(池田等人,2017年)。
除了与采血蝇和跳蚤的互动外,蝙蝠与其他外寄生虫的关联可能解释这种多病原体感染。在巴西,巴尔通体已在19.8%(40/202)的虻科苍蝇中被检测到(Amaral等人,2018年)。
使用MEGA12软件,采用Tamura 3参数模型和1000次自助法复制来评估分支支持。以锥虫作为外群。
我们的狂犬病病毒序列在系统发育树上的位置突显了蝙蝠在狂犬病病毒传播链中的重要性,特别是在不同哺乳动物物种共享栖息地的地方。蝙蝠狂犬病病毒序列与人类和野生犬科动物序列之间的系统发育接近性表明可能存在流行病学联系,加强了狂犬病病毒潜在宿主和宿主的多样性。
本研究的一个重要局限性是只有一只蝙蝠显示出共感染,这可能限制了结果的普遍性。同时,这一发现是相关的,因为它突显了蝙蝠中病原体之间相互作用的复杂性及其作为多个宿主的潜在能力。此外,将这一发现在巴西监测系统中进行情境化是很重要的,该系统优先考虑收集和分析发现的坠落或处于异常情况的蝙蝠,例如在室内。尽管这个系统在检测狂犬病例方面很有效,但可能低估了其他人畜共患病的情况,因为对其他病原体在蝙蝠中的主动监测仍然有限。
结论
本研究报告首次在南美洲东南部的一只黑蝙蝠体内发现了狂犬病毒、贝氏巴尔通体和李氏利什曼原虫的共感染。研究结果强调了这些动物作为具有不同传播周期的多种人畜共患病原体的潜在多重宿主的潜力,加强了对人类、家畜和野生动物共存区域的综合性监测需求。在单一宿主体内检测到这些病原体,突显了在人工化环境中病原体与宿主相互作用的复杂性,以及进一步研究评估这些共感染对生态和流行病学影响的重要性。
作者贡献 所有作者均参与研究的构思与设计。材料准备和数据收集由Danilo Alves de Franca、Benedito D. Menozzi和Helio Langoni完成。方法和结果分析由Danilo Alves de Franca和Bruna L. D. Ribeiro进行。稿件初稿由Danilo Alves de Franca撰写。所有作者阅读了稿件,提供了写作意见并批准了最终稿件。
资助 本项工作得到了圣保罗州研究基金会(授权编号2022/07124-6和2022/10976-4)的支持。
数据可用性 在当前研究期间没有生成或分析任何数据集。
声明
伦理审批 动物使用伦理委员会(编号0259/2022),巴西圣卡塔琳娜州立大学兽医学院及动物技术系。环境许可(编号85973-1)。
利益冲突 作者没有需要声明利益冲突的内容。
Hits: 78
- 狂犬病(2025年6月更新)
- 狂犬病 你应该知道的
- 预防和治疗狂犬病的狂犬病疫苗
- 狂犬病可能成为历史
- 看看每种疫苗:狂犬病疫苗
- 阿拉巴马州妇女在从车上卸下杂货时被狂犬病狐狸咬伤:“公共卫生威胁”
- 应对狂犬病
- 全球疫苗免疫联盟将促进全球疫苗免疫联盟、世卫组织和阿联酋50多个国家获得拯救生命的人类狂犬病疫苗
- 美国CDC发布了将狗带入美国的新规定,旨在防止狂犬病
- 全国狂犬病监测方案
- 2024年东帝汶首例狂犬病确诊病例
- WHO实况报道 狂犬病
- 狂犬病
- 狂犬病疫苗接种:给医疗保健提供者的信息
- 特定感染的流行病学和控制:狂犬病
- 狂犬病疫苗接种的注意事项或禁忌症
- ACIP 使用减少(4剂)疫苗计划进行暴露后预防以预防人类狂犬病
- 实施良好的狂犬病暴露后预防方案
- ACIP 使用修改后的暴露前预防疫苗接种计划预防人类狂犬病,美国,2022年
- 西班牙犬类狂犬病免疫保护的影响因素
- 减少犬的进口等待期-对狂犬病恐惧科学合理的变化
- 老年人对狂犬病暴露后预防无应答:挑战和方法
- 狂犬病疫苗的效力、免疫原性和有效性:体外和体内方法
- 狂犬病人免疫球蛋白 (HRIG)
- “震惊”的妇女,超过6,000美元的救命狂犬病治疗账单
- 负鼠感染狂犬病毒对这种病毒在城市环境中的传播敲响了警钟
- 索马里蜜獾感染未报告的首例疑似人狂犬病病例研究
- 两名狂犬病研究人员解释如何保护自己免受这种疾病的侵害
- 临床医生常见的狂犬病问题
- 狂犬病疫苗
- 世界卫生组织关于狂犬病的常见问题
- 狂犬病实况报道
- 狂犬病
- 狂犬病是一种古老的、不可预测的、可能致命的疾病——两位狂犬病研究人员解释了如何保护自己
- 狂犬病疫苗短缺:给卫生专业人员的建议
- 7例被狂犬咬伤者接种狂犬病疫苗的效果观察
- 全球狂犬病风险:狂犬病疫苗短缺期间的实用建议
- 巴西一个土著村庄被蝙蝠掠夺的高风险:警告狂犬病在马卡利人中重新出现
- 评估在澳大利亚蝙蝠狂犬病病毒和狂犬病暴露后预防中血清学的益处
- 蝙蝠处理者的狂犬病暴露前预防
- 基于狂犬病中和抗体水平的狂犬病加强剂时机指南
- 施用狂犬病疫苗和免疫球蛋白
- 为狂犬病暴露后预防开发的单克隆抗体鸡尾酒疗法
- 狂犬病预防处置常见问题解答(成大生物)
- 世界卫生组织狂犬病专家磋商会 第三版报告
- 世卫组织对狂犬病的看法
- 新型狂犬病疫苗
- 狂犬病病毒可以抗强大的天然免疫
- 狂犬病的被动免疫
- 亚洲狂犬病预防
- 秘鲁高风险人群的大规模狂犬病暴露前预防
- 人用狂犬病疫苗
- 野生动物狂犬病疫苗
- 狂犬病和狂犬病疫苗的历史
- 狂犬病的生命周期、传播和发病机制
- 狂犬病暴露后管理指南(英国 2023年1月)
- 2022年印度马哈拉施特拉邦一名幼儿出现临床狂犬病症状后存活
- 第27章:狂犬病
- 狂犬病:问题和答案
- 《狂犬病暴露预防处置工作规范(2023年版)》解读问答
- 狂犬病暴露预防处置工作规范(2023年版)
- 狂犬病
- 狂犬病相关问题解答
- 疾病控制中心加强了对进口狗的狂犬病管理,原因如下
- 关于狂犬病
- 一名加州艺术教师在教室遭遇蝙蝠后死于狂犬病
- WHO 实况报道 动物咬伤 2024年1月
- 疫苗接种和免疫 狂犬病
- 世卫组织推荐皮内途径用于狂犬病暴露后预防
- 狂犬病疫苗:世卫组织立场文件–2018年4月
- WHO实况报道 狂犬病 2024年6月
- 狂犬病-结束忽视, 实现可持续发展目标 被忽视的热带病路线图 2021-2030
- 动物狂犬病
- 宠物主人对疫苗的犹豫越来越多——一位公共卫生专家解释了为什么这很重要
- 兽用狂犬病疫苗
- 何时寻求狂犬病医疗护理
- 猫
- 关于获得人类狂犬病疫苗的社会正义观点
- 评估新的狂犬病暴露后预防(PEP)方案或疫苗
- 卫生和福利部-狂犬病
- 狂犬病暴露后预防用单克隆抗体开发的最新进展:临床开发管线现状综述
- 世卫组织对狂犬病免疫接种的立场 – 2018 年最新情况
- 恐水症:1874 年的狂犬病和改变一切的救命疫苗
- 新的人类狂犬病疫苗正在研发中
- 为什么消除狂犬病?
- 为什么现在要根除狂犬病?
- 狂犬病-NFID
- 为狗接种疫苗 – 它们不会因接种疫苗而患上自闭症
- 宠物主人版 狗的狂犬病
- 猫的狂犬病
- 狂犬病-国际脑炎协会
- 器官移植后如何感染狂犬病?
- 器官移植患者感染狂犬病后死亡
- 人类狂犬病疾病负担:Rabipur®疫苗的潜在预防作用
- 印度东北部农村的狂犬病:强调同一健康方法紧迫性的病例报告
- 狂犬病毒糖蛋白的深度突变扫描确定了突变限制和抗体逃逸突变
- 人类狂犬病:一种治疗方法
- 通过受感染狗唾液污染的隐形眼镜传播狂犬病病毒(病例报告)
- 越南暴露前预防计划中基于灭活 vero 细胞的狂犬病疫苗 (Rabivax-S) 的安全性和免疫原性
- 狂犬病生物制剂在 III 类动物暴露儿科患者中的安全性和临床疗效的前瞻性研究
- 重复狂犬病疫苗接种揭示人类中和抗体的分子趋同
- 狂犬病病毒的感染和预防
- 关于 Zagreb 或 Essen 方案后加强疫苗接种的免疫原性和安全性的 3 期临床试验
- 通过 RABV-G 结构域筛选和异源初免-加强免疫优化狂犬病 mRNA 疫苗功效
- 狂犬病实验室诊断的最新情况
- 三色蝠相关狂犬病毒引起的一例人狂犬病的报道
- 一种核苷修饰的狂犬病mRNA疫苗在小鼠和非人灵长类动物中诱导出持久而全面的免疫反应
- 狂犬病的识别和治疗:病例报告和讨论
- 人类狂犬病的诊断困难:病例报告和文献回顾
- 狂犬病感染的边缘系统症状
- 狂犬病脑炎的生存率
- 人类狂犬病:2016年更新
- 突尼斯一名狂犬病幸存儿童:病例报告
- 肯尼亚蝙蝠携带的Duvenhage病毒导致人类狂犬病死亡:昏迷诱导、氯胺酮和抗病毒药物治疗无效
- 病例报告:狂犬病幸存者长期随访中的神经功能恢复与血清学反应
- 蝙蝠和食肉动物中狂犬病的比较病理学及其对人类溢出影响
- 病例报告:狂犬病生存者:印度病例系列
- 评估人类抗狂犬病疫苗的反应:一种快速、高通量适应性、基于假病毒的中和试验,作为快速荧光病灶抑制试验 (RFFIT) 的替代方案
- 病例报告:狂犬病幸存者长期随访中的神经功能恢复与血清学反应
- 巴西巴伊亚州狂犬病爆发的流行病学与分子分析
- 俄罗斯下诺夫哥罗德地区动物狂犬病(2012-2024年):描述性与预测性流行病学分析
- 巴西东南部一只黑鼠耳蝠(黑鼠耳蝠属物种Myotis nigricans)感染狂犬病病毒、科勒巴尔通体菌以及婴儿利什曼原虫的混合感染情况
- 2013-2023 年悉尼大都会居民狂犬病暴露后治疗:回顾性病例系列分析
- 用肌肉靶向脂质纳米颗粒配制的狂犬病病毒 G 蛋白 mRNA 在小鼠体内的免疫原性
- 关于狂犬病的 One Health Lens:马拉维利隆圭农村狂犬病病毒的人类-蝙蝠相互作用和基因组见解
- 阿根廷低地貘 (Tapirus terrestris) 麻痹狂犬病的首次报告
- 由表达狂犬病糖蛋白的非复制型病毒载体痘苗病毒天坛株改良株(MVA)和腺病毒5型(Ad5)所诱导产生的对狂犬病的保护作用
- 印度野生猫科动物中狂犬病毒的免疫病理与分子筛查:揭示从野外到城市的溢出效应
- 韩国貉 (Nyctereutes procynoide koreensis) 接种了含有 ERAGS-GFP 毒株的新型狂犬病诱饵疫苗的保护性抗体反应
- 宠物护理 疫苗
- 为您的宠物接种疫苗
- 狂犬病感染:如何保护您的孩子
- 新型狂犬病暴露后预防单克隆抗体鸡尾酒疗法的安全性和耐受性
- “愤怒的狂犬病”人类图画案例
- 蝙蝠咬伤
- 研究蝙蝠病毒如何帮助预防人畜共患疾病
- 狂犬病:症状繁多、宿主广泛的恶魔病毒——罗德尼·罗德解读
- 狂犬病的同一健康:不仅适用于动物
- 狂犬病和蛔虫:浣熊相关传染病
- 丽莎病毒 很罕见,但致命。如果蝙蝠咬你怎么办?
- 海地在四个省开展新的疫苗接种运动,加强防治犬类狂犬病
- 家养及其他动物狂犬病病毒感染的挑战:病毒学层面的困惑、惊愕、懊恼与好奇交织
- 双重战斗:狂犬病和蛇咬伤在东南亚的毒害
- 狂犬病在城市无人管理的猫群中爆发 — 马里兰州,2024 年 8 月
- 尼泊尔正力争在2030年前消灭狂犬病,但领导人表示,现在是加大疫苗投资的时候了。
- 现代狂犬病疫苗的诞生历程
- 一种基于mRNA的新型狂犬病病毒多表位疫苗:通过反向疫苗学和免疫信息学进行计算机设计
- 快速荧光灶抑制试验(RFFIT)的验证与评价 —— 作为狂犬病疫苗相对效价间接检测方法的中和抗体定量检测:NIH 试验的替代方案
- 暴露后狂犬病预防失败案例研究
- 2021年明尼苏达州一起经确认的人畜共患病暴露后,疑似因宿主介导导致暴露后预防失败的致命人类狂犬病感染病例
- 给野生动物投喂疫苗:口服狂犬病疫苗的故事
- 降低肯尼亚西部臭名昭著的狗市狂犬病风险
- 印度的狂犬病
- 海地的狂犬病
- 2024年10月至2025年2月,美国发生通过实体器官移植传播狂犬病的病例,该器官移植供体未确诊患有狂犬病
- 两例死亡与狂犬病感染的肾移植有关
- 明尼苏达州一家奶牛场的牛群中爆发狂犬病聚集性疫情——2024年
- 摩洛哥的狂犬病
- 2024年肯塔基州和俄亥俄州输入性人类狂犬病病例
- 2024年明尼苏达州和加利福尼亚州人类狂犬病死亡人数
- 反疫苗人士对宠物狂犬病 RNA 疫苗的虚假宣传
- 新型口服狂犬疫苗能否帮助我们最终终结这种疾病?
- NACI 2026 年 1 月 30 日发布的声明摘要:狂犬疫苗用于暴露前预防 (PrEP) 的最新指南
- 狂犬疫苗的给药途径:数据比较
- 猫会得狂犬病吗?
- 2022-2024年中欧狂犬病疫情再度出现
- 研讨会报告:改进狂犬病血清学方法和程序的标准化
- 我们来谈谈狂犬病检测