
Contents
- 1 引言
- 2 结果
- 3 狂犬病mRNA疫苗在小鼠中诱导出强烈的抗狂犬病毒(RABV)特异性抗体
- 4 狂犬病mRNA疫苗能在小鼠中诱导强烈的细胞免疫反应
- 5 狂犬病mRNA疫苗在小鼠中诱导了持续的体液免疫反应
- 6 狂犬病mRNA疫苗在恒河猴中诱导了强烈的免疫应答
- 7 狂犬病mRNA疫苗在恒河猴中显示出良好的安全性
- 8 讨论
- 9 材料与方法
- 10 病毒攻击
- 11 样本采集和处理
- 12 流式细胞术
- 13 AIM+T细胞
- 14 免疫染色法
- 15 浆母细胞和生发中心B细胞
- 16 酶联免疫斑点试验
- 17 血浆细胞因子的定量
- 18 统计分析
- 19 数据和代码可用性
- 20 致谢
- 21 作者贡献
- 22 利益声明
- 23 生成式AI和AI辅助技术在写作过程中的应用声明
- 24 补充信息
- 25 参考文献
doi: 10.1016/j.ymthe.2024.12.041. Epub 2024 Dec 31.
A nucleoside-modified rabies mRNA vaccine induces long-lasting and comprehensive immune responses in mice and non-human primates
- PMID: 39741409
狂犬病是一种致死性的人畜共患传染病。针对狂犬病毒的疫苗显著减少了该病的死亡人数。然而,所有获批的狂犬病疫苗均为灭活疫苗,其免疫原性有限且接种程序复杂。迫切需要一种能提供持续全面保护的新型疫苗。在这里,我们开发了一种新型狂犬病mRNA疫苗候选物,其中包含序列优化后的mRNA,编码全长糖蛋白,这些糖蛋白被包裹在可离子化的脂质纳米颗粒中。在小鼠和恒河猴中,狂犬病mRNA疫苗的免疫原性优于获批疫苗,尤其是在诱导持久的中和抗体和记忆B细胞方面。小鼠单次接种1.5微克mRNA疫苗可提供完全保护,抵御致死性的狂犬病病毒挑战。此外,mRNA疫苗能强烈激活细胞免疫反应,并适度释放多种细胞因子。总之,我们的数据表明,狂犬病mRNA疫苗在小鼠和恒河猴中的表现均优于获批的灭活疫苗。这突显了mRNA平台在开发下一代狂犬病疫苗方面的潜力。
确保预防。这一复杂的免疫接种方案给农村和高风险人群带来了负担。每年约有6万人死于狂犬病。这些数据促使人们开发一种新型狂犬病疫苗,能够提供持久和全面的保护。
mRNA平台在抗击COVID-19中发挥了至关重要的作用。mRNA疫苗的生产过程简单、安全且产量高。此外,mRNA疫苗能有效介导细胞免疫反应和生发中心(GC)的形成。GC对长寿命浆细胞(LLPCs)和记忆B细胞(MBCs)的产生有促进作用,有助于形成更全面和持久的免疫反应,这可能会弥补灭活疫苗的局限性。近期,德国图宾根的CureVac AG公司开发了一种未经修饰的狂犬病mRNA疫苗,该疫苗耐受性良好,并且在低剂量下能够引发保护性的中和抗体反应。然而,在一项Ⅰ期试验中,高剂量诱导了不可接受的反应原性。使用修饰的核苷酸可以提高mRNA的稳定性、蛋白质翻译效率,并降低反应原性。
引言
狂犬病是由狂犬病病毒(RABV)引起的一种人兽共患传染病。已在全球150多个国家和地区被发现。RABV是一种包膜的单链RNA病毒,具有五种结构蛋白,其中糖蛋白(G)能够诱导特异的中和抗体。当人类被感染动物抓伤或咬伤时,RABV逐渐侵入神经系统,导致严重的神经炎症和接近100%的死亡率。幸运的是,这种致命疾病可以通过疫苗接种来预防。然而,目前唯一获得许可的狂犬病疫苗是灭活疫苗,其免疫原性有限。世界卫生组织(WHO)推荐接种三剂用于暴露前预防,四至五剂用于暴露后预防。
收稿日期:2024年5月16日;接受日期:2024年12月27日
https://doi.org/10.1016/j.ymthe.2024.12.041。
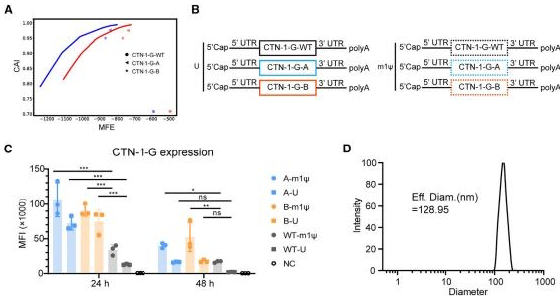
图1.狂犬病mRNA疫苗的构建和表征
(A)CTN-1-G序列的密码子优化。未优化的序列表示为CTN-1-G-WT,两个优化后的序列分别表示为CTN-1-G-A/B。(B)CTN-1-G mRNA结构的示意图。U,尿苷;ml,Nl-甲基-假尿苷。(C)体外表达。2μg的CTN-1-G mRNA被转染到4×10^5个HEK293T细胞中。通过流式细胞术在转染后24小时和48小时检测G蛋白的表达。(D)LNP-CTN-1-G-mRNA颗粒大小的代表性分布图。实验独立重复三次。数据以平均值±标准差的形式呈现。显著性差异使用单因素方差分析(p<0.05,p<0.01,和p<0.001;ns,无显著性差异)进行分析。
mRNA疫苗,并评估其在啮齿类动物和犬中的效力。33-35 进一步评估疫苗的有效性并在非人灵长类动物中进行深入的疫苗反应特征描述是必要的。
在本研究中,我们设计了一种新型的含有线性设计算法优化的mRNA疫苗,这些mRNA被包裹在可电离的脂质纳米粒子(LNP)配方中。线性设计算法同时考虑了最小自由能(MFE)和密码子适应指数(CAI),从而实现了更稳定和高效的序列优化。我们进一步与恒河猴和小鼠中的灭活疫苗进行了并排比较。我们的狂犬病mRNA疫苗能够在低剂量下在恒河猴中诱导高水平的病毒中和抗体(VNAs)以及持续的体液和细胞免疫应答和免疫记忆,且具有良好的安全性。单次接种1.5微克mRNA疫苗能够在小鼠中提供对抗致死性狂犬病病毒(RABV)挑战的完全保护。这些发现可能增进我们对狂犬病mRNA疫苗作用机制的理解,并指导未来N1-甲基-假尿嘧啶核苷(m1w)修饰的应用。
结果
体外设计并构建编码CTN-1-G蛋白的狂犬病mRNA疫苗
G蛋白是狂犬病的主导免疫原,已被广泛用作疫苗开发的关键抗原候选。这里,我们选择了CTN-1株的G蛋白序列(GenBank:FJ959397.1)作为抗原,该序列与中国街头的RABV株在进化上相关。使用线性设计算法,我们设计了两个优化序列(CTN-1-G-A/B),与野生型序列(CTN-1-G-WT)相比,具有更高的密码子适应指数(CAI)和更低的自由能(MFE)(图1A)。优化的mRNA通过体外转录(IVT)合成,并掺入尿嘧啶(U)或m1w(图1B)。所有候选序列在转染至HEK293T细胞后都产生了强大的蛋白质表达。密码子优化和m1u修饰进一步提高了表达效率,并延长了G蛋白的半衰期(图1C)。候选mRNA疫苗配方通过将CTN-1-G-mRNAs包封进脂质纳米颗粒(LNP)递送系统中制备。动态光散射(DLS)分析证实,LNP-mRNA的粒径为120-130纳米(图1D),据报道其具有高度免疫原性。40
狂犬病mRNA疫苗在小鼠中诱导出强烈的抗狂犬病毒(RABV)特异性抗体
候选mRNA疫苗的免疫原性首先在与商业用灭活疫苗的比较中,在BALB/c小鼠中进行评估。我们在第0天和第21天用递增剂量的mRNA疫苗(0.5、1.5或5微克)或灭活疫苗(0.25IU)免疫小鼠,并在之后确定抗体反应(图2A)。候选mRNA疫苗能够产生剂量依赖的人体免疫反应,如在第14天、第28天和第42天的特异性结合抗体所示(图2B-2D)。具体而言,CTN-1-G-A和CTN-1-G-B在诱导抗体反应方面整体能力相当,显著强于灭活组(除第14天低剂量组外,p<0.05)。平均抗体滴度
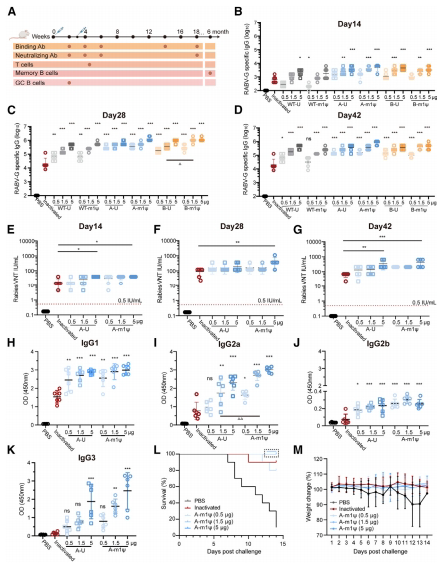
图2.狂犬病mRNA疫苗诱导小鼠产生强大的体液免疫应答
(A)实验设计。(B-K)用0.5、1.5或5微克狂犬病mRNA、灭活疫苗(0.25IU)或PBS在第0天和第21天肌内免疫6只BALB/c小鼠,收集血清以检测结合抗体和病毒中和抗体(VNA)滴度。使用酶联免疫吸附试验(ELISA)在第14天(B)、28天(C)和42天(D)测定特异性总IgG滴度。采用荧光灶抑制中和试验(FAVN)在第14天(E)、28天(F)和42天(G)滴定VNA。使用ELISA在第28天分析包括IgG1(H)、IgG2a(I)、IgG2b(J)和IgG3(K)在内的热型IgG。(L和M)10只BALBic小鼠接受单剂量CTN-1-A-m1、灭活疫苗(0.25IU)、PBS疫苗接种,然后在14天后用致死剂量的狂犬病毒进行挑战。监测14天的存活率(L)和体重减轻(M)。数据以平均值±标准差表示。使用单因素方差分析(ANOVA)分析显著性差异(mRNA组与灭活组比较,p<0.05,p<0.01,p<0.001;相同剂量下U组与mlp组比较, p<0.05, p<0.01;ns,无显著性差异)。
免疫后未观察到小鼠体重的显著差异(图S1)。我们进一步表征了免疫球蛋白G(IgG)亚型,并发现mRNA疫苗能够以剂量依赖的方式在BALB/c小鼠中诱导更高水平的特异性IgG1、IgG2(主要是IgG2a)和IgG3抗体(图2H-2K)。
为了探索体内保护效果,我们进行了致死性狂犬病病毒(RABV)挑战实验。在初次免疫后,mRNA疫苗诱导的病毒中和抗体(VNA)高于保护阈值(图2E),因此我们在这里采用了单次免疫策略。如图2L所示,CTN-1-G-A组中1.5μg的CTN-1-G-A-m1μ的VNA略高于CTN-1-G-B组;因此,我们在后续分析中重点关注CTN-1-G-A疫苗。我们测定了VNA滴度,它与保护效率呈正相关(图2E-2G)。接受5μg剂量的CTN-1-G-A-m1μ的小鼠在初次免疫和加强免疫后产生的VNA滴度最高,显著高于灭活组(图2E,p<0.05;图2F,p<0.01;图2G,p<0.001)。第14天和第28天的平均VNA滴度分别为40.5和445.6IU/毫升,而相应滴度在灭活组中为16.5和91.65IU/毫升。0.5μg CTN-1-G-A mRNA诱导的VNA滴度最低,但仍与灭活组相当。与尿苷相比,m1w掺入的疫苗产生的VNA滴度略高,但没有统计学差异。值得注意的是,所有疫苗组的小鼠在初次免疫后14天均产生了高于0.5IU/毫升的VNA滴度。接种狂犬病mRNA疫苗的小鼠获得了针对狂犬病毒的完全保护,而灭活疫苗组的一只小鼠在挑战后10天死亡。挑战后未观察到小鼠体重的显著差异(图2M)。总体而言,上述结果证明了狂犬病mRNA疫苗能在小鼠中诱导出强烈的体液免疫反应,并且单次接种1.5μg CTN-1-G-A-m1μmRNA就能提供完全保护。
狂犬病mRNA疫苗能在小鼠中诱导强烈的细胞免疫反应
由于T细胞在调节体液免疫和抵御病原体方面起着关键作用,我们进一步研究了mRNA疫苗诱导的T细胞反应。T细胞激活诱导标记(AIM)实验显示,G特异性AIM+ CD4+ T细胞(图3A和3B,p<0.01)和AIM+ CD8+ T细胞(图3C)的诱导作用更强。
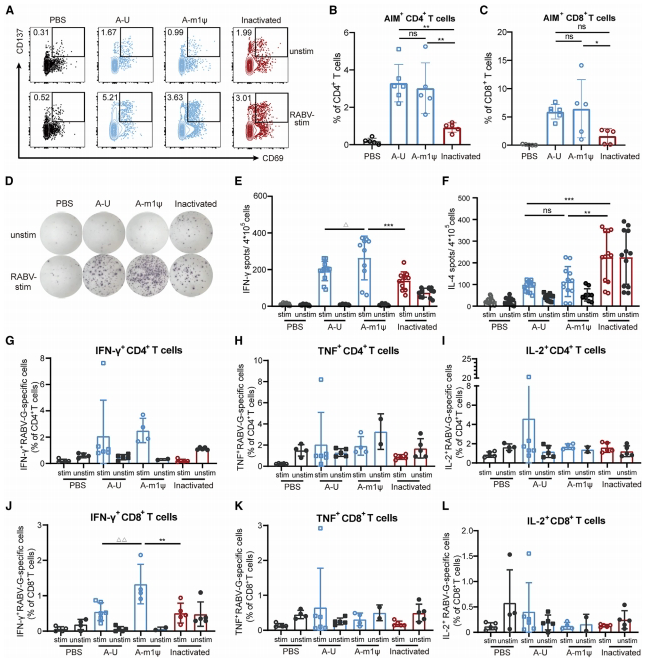
图3.狂犬病mRNA疫苗在小鼠中诱导的细胞免疫应答比已获许可的灭活疫苗更强
在抗原刺激下,CTN-1-G-A mRNA疫苗接种的小鼠脾细胞中,与对照组相比(p<0.05),我们发现CTN-1-G-A mRNA免疫能诱导出更高水平的干扰素-γ(IFN-γ)分泌型而非白细胞介素-4(IL-4)分泌型的T细胞,这一结果显著高于灭活疫苗(图3D-3F和S3B),表明其引发的是偏向Th1(T辅助1型)的免疫反应。最后,我们进行了胞内细胞因子染色(ICS)实验,以评估特异性针对G蛋白的IFN-γ、肿瘤坏死因子(TNF)、IL-2分泌型T细胞的频率(图3G-3L,S2以及S3C-S3H)。结果显示,与灭活疫苗相比,接种了CTN-1-G-A m1u修饰mRNA的小鼠CD4+(图3G和S3C)和分泌IFN-γ的CD8+ T细胞(图3J和S3F,p<0.01)频率更高。在TNF(图3H、3K、S3D和S3G)或IL-2(图3L、3L、S3E和S3H)方面未观察到显著差异。所有这些数据均证明,与小鼠体内灭活的疫苗相比,CTN-1-G-A mRNA在诱导偏向Th1的G蛋白特异性T细胞反应方面更为有效,而m1u修饰进一步增强了T细胞的活化作用。
狂犬病mRNA疫苗在小鼠中诱导了持续的体液免疫反应
为了评估长期保护效果,我们在第98天和第126天测量了特异性IgG滴度(图4A和4B),以及在第126天的病毒中和抗体(VNA)滴度(图4Q)。CTN-1-G-A m1w组的平均IgG滴度在加强免疫后15周仍然显著高于灭活组(图4B,p<0.001)。接受0.5、1.分别给予5μg剂量的CTN-1-G-A-m1μ mRNA,而灭活组的平均病毒核糖核酸(VNA)滴度降至28.6IU每毫升(图4C)。
在遇到特定抗原后,浆细胞可以迅速增殖并分化成浆细胞,这对于长期保护至关重要。为了评估CTN-1-G-A-m1μ mRNA是否能诱导记忆免疫,我们在免疫后6个月通过分析CD19+ B220+ FAS-IgD+ CD38+ CTN-1-G+细胞的丰度来检查浆细胞(图4D、4E和S4A)。与接种灭活疫苗的小鼠相比,接种CTN-1-G-A mRNA的小鼠观察到显著更多的浆细胞(p< 0.001),而在灭活疫苗组几乎检测不到浆细胞的比例。由于生发中心反应在产生长寿命浆细胞和浆细胞方面至关重要,进一步调查了CTN-1-G-A mRNA在诱导生发中心B细胞中的作用。流式细胞术结果显示,mRNA疫苗诱导的抗原特异性生发中心B细胞水平显著高于灭活疫苗(p< 0.05),而总生发中心B细胞的频率没有显著差异(图4F-4I和S4B)。综合这些数据表明,CTN-1-G-A mRNA在小鼠体内诱导了持续的体液免疫应答,这通过抗体滴度、浆细胞频率和生发中心反应得到了证实。
狂犬病mRNA疫苗在恒河猴中诱导了强烈的免疫应答
非人灵长类动物是临床疫苗评估的理想动物模型,其中核苷酸修饰的狂犬病mRNA疫苗诱导的免疫应答的质量和特性大多仍不为人所知。因此,我们评估了由CTN-1-G-A-m1w mRNA(30微克)和灭活疫苗(2.5IU)在恒河猴中诱导的特异性抗体和B细胞反应、T细胞反应以及炎症细胞因子的释放(图5A)。与我们在小鼠模型中的发现一致(图2),CTN-1-G-A-m1w mRNA诱导的非人灵长类动物中的IgG和VNA滴度高于灭活疫苗(图5B和5C)。初次免疫后14天,mRNA组的VNA滴度比灭活疫苗组的23.4IU/毫升高出42.9IU/毫升,远高于保护阈值(>0.5IU/毫升)。加强免疫后1周,VNA滴度达到峰值,之后逐渐下降。值得注意的是,免疫后27周,mRNA组和灭活疫苗组的VNA滴度分别为94.5IU/毫升和6.5IU/毫升,与各自峰值相比下降了4.08倍至17.99倍(图5C,p<0.05)。mRNA组在免疫后19周的特异性类转换IgG+浆母细胞的频率也更高(图5D、5E和S5A),证明了CTN-1-G-A-m1w的长期保护效率。
鉴于在小鼠模型中mRNA疫苗强烈诱导了Th1型T细胞,我们在恒河猴中进行了ICS分析以验证这一结论(图S5B)。与之前的结果一致,mRNA疫苗诱导了更高频率的产生IFN-γ而非TNF-α或IL-2的T细胞,相较于灭活疫苗(图5F-5H和S6)。
为了初步评估疫苗接种对先天免疫的激活作用,我们量化了恒河猴中的炎性细胞因子(图5I和S7)。检测到的多数细胞因子在接种后1天出现短暂升高,并在1-2周内恢复到正常水平,表明mRNA疫苗刺激的细胞因子分泌是可控的。值得注意的是,我们在mRNA组的两次免疫后都观察到IL-6显著增加,而IL-6已被报告为形成生发中心的关键细胞因子。综上所述,我们证明了CTN-1-G-A-m1μ在恒河猴中诱导的免疫反应比灭活疫苗更为强烈,其特征为持续较长时间的病毒中和抗体、更多的记忆B细胞以及强大的偏向Th1的T细胞反应。
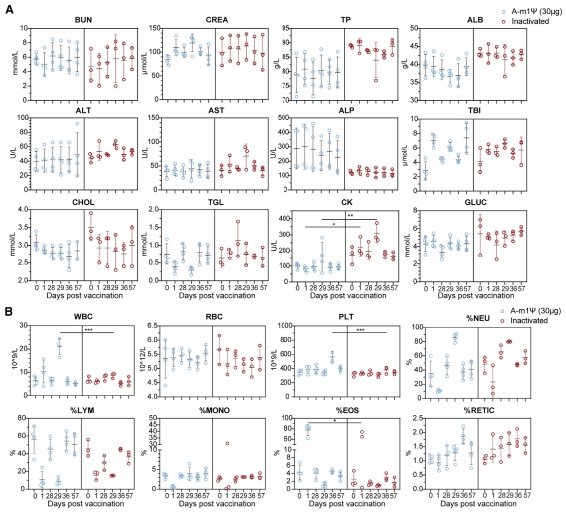
狂犬病mRNA疫苗在恒河猴中显示出良好的安全性
最后,我们对恒河猴中的狂犬病mRNA进行了初步的安全性评估。我们监测了接种疫苗前后血清中的生化参数变化(图6A)。结果表明,恒河猴能够很好地耐受这种核苷酸修饰的狂犬病mRNA疫苗,没有肝或肾毒性。血液生化参数的波动在1周内恢复到基线水平。值得注意的是,mRNA疫苗组在初次和加强免疫后的肌酸激酶(CK)水平较低(p<0.05, p<0.01),这暗示较低的心脏毒性。此外,我们还评估了血液学参数(图6B)。免疫接种后,白细胞计数、血小板计数和嗜酸性粒细胞百分比出现短暂升高,并在7天内恢复正常。总体而言,我们的数据表明CTN-1-G-A-m1μmRNA疫苗在非人灵长类动物中具有良好安全性。
讨论
狂犬病是一种广泛传播的致命人畜共患病,对公共卫生构成巨大威胁。疫苗接种是预防狂犬病的最具成本效益的策略。
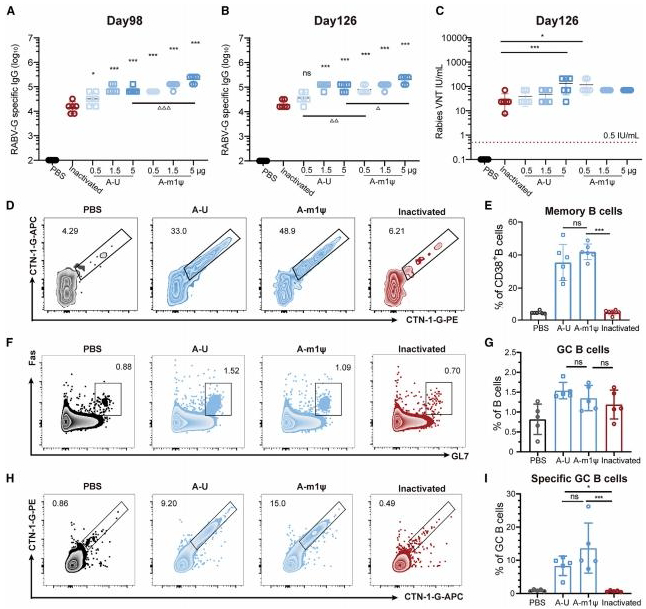
图4.狂犬病mRNA疫苗诱导小鼠产生高水平的持续性体液免疫
(A-C) 用图2A所述方法对6只BALB/c小鼠进行免疫。在第98天(A)和第126天(B),通过酶联免疫吸附试验(ELISA)检测血清中特异性IgG滴度。(C) 在第126天对病毒中和抗体(VNA)进行滴定。(D和E) 在第0天和第21天,用5微克狂犬病mRNA疫苗、0.25IU灭活疫苗或磷酸盐缓冲液(PBS)对6只BALB/c小鼠进行免疫。在初免后六个月,分析外周血单个核细胞(PBMC)中的记忆B细胞(MBC)。(D) 记忆B细胞的代表性流式细胞术图。(E) 记忆B细胞的频率。(F-I) 用5微克狂犬病mRNA疫苗、0.25IU灭活疫苗或磷酸盐缓冲液对5只BALB/c小鼠进行免疫。在免疫后14天,评估脾脏中的生发中心B细胞(GC B cells)。(F) 总生发中心B细胞的代表性流式细胞术图。(G) 总生发中心B细胞的频率。(H) 针对CTN-1-G的特异性生发中心B细胞的代表性流式细胞术图。(I) 针对CTN-1-G的特异性生发中心B细胞的频率。数据以平均值±标准差的形式呈现。使用单因素方差分析(ANOVA)分析mRNA组与灭活疫苗组之间的显著差异(p<0.05和**p<0.001;ms,无显著性差异);使用单因素方差分析比较同一剂量下U组与mlq组之间的显著差异(p<0.05、p<0.01和***p<0.001;ms,无显著性差异)。
对照组。然而,所有获批的狂犬病疫苗都是灭活疫苗,其免疫原性有限且需要多次注射。10,11,49 普遍认为mRNA疫苗在诱导T细胞和B细胞反应方面效力强劲。50,51 COVID-19的mRNA疫苗数据也显示出了比灭活疫苗更高的保护效率。因此,近年来已经研发了几种狂犬病mRNA疫苗,并在啮齿类动物和犬中评估了其有效性和安全性。在本研究中,我们
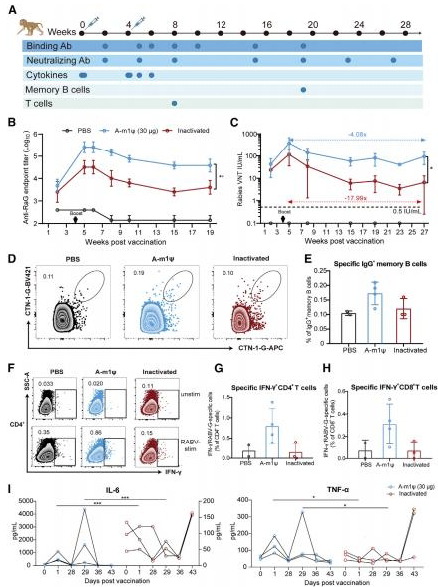
图5. 狂犬病mRNA疫苗在恒河猴中诱导了强烈的免疫反应
(A)恒河猴的实验设计。恒河猴通过肌内注射接种了30微克CTN-1-G-A、2.5IU获批的灭活狂犬病疫苗或磷酸盐缓冲液(PBS)。血清和外周血单个核细胞按计划采集。(B)进行酶联免疫吸附试验以测量特异性IgG滴度。(C)执行荧光抗狂犬病毒中和试验以评估病毒中和抗体(VNA)滴度。(D)免疫后19周,外周血单个核细胞中IgG+记忆B细胞(CD3- CD14- CD20- IgM- IgG+ CTN-1-G+)的代表性流式细胞术图。(E)IgG记忆B细胞的频率。(F)加强免疫后4周,外周血单个核细胞中分泌干扰素-γ的CD4 T细胞的代表性流式细胞术图。通过细胞内染色法分析干扰素-γ CD4+ T细胞(G)和干扰素-γ+ CD8+ T细胞(H)的频率。(I)血清细胞因子的定量检测。数据表示为均值±标准差。使用单因素方差分析或非配对t检验分析显著差异(p<0.05,p<0.01,和p<0.001)。
单独5微克剂量组在14天后诱导出约40 IU/毫升的VNA滴度。我们假设线性设计算法的应用有助于提高翻译效率和疫苗免疫原性,因此,CTN-1-G-A-m1μ疫苗优于其他已报告的疫苗。在恒河猴模型中,VNA滴度在加强免疫后1周达到峰值(385.8 IU/毫升),并在27周后下降了4.08倍至94.5 IU/毫升,而灭活组的VNA滴度在17.99倍下降至6.5 IU/毫升。随着外周血单个核细胞(PBMCs)中微小抱子菌(MBCs)频率的增加,我们证实CTN-1-G-A-m1能在非人灵长类动物中产生比灭活疫苗更强大、更持久的抗体和B细胞反应。值得注意的是,我们采用了0-28天的免疫接种计划,以展示疫苗反应的质量和特点。该计划可能不是用于暴露后治疗的最佳方案,需要进一步研究。我们发现,在小鼠和恒河猴模型中,我们的狂犬病mRNA疫苗能诱导出比商业用灭活疫苗显著更强的抗体反应。此外,在CTN-1-G-A-m1q组中的病毒中和抗体(VNA)滴度甚至高于先前的报告。在小鼠模型中,与接种后14天产生约10-15IU每毫升(IU/mL)VNA滴度的单剂量5微克mRNA免疫相比,我们的疫苗诱导了更强的反应。
对于狂犬病mRNA疫苗诱导的强大而持久的疫苗反应背后的作用机制仍知之甚少,我们进行了进一步的研究。我们证明了mRNA疫苗能够诱导更多的抗原特异性生发中心B细胞,这对于产生记忆B细胞和长期抗体至关重要。同时,我们在恒河猴模型中发现在免疫后第1天IL-6短暂升高,其他用脂质纳米颗粒包封的基于mRNA的疫苗也有类似报道。IL-6是与生发中心反应相关的关键细胞因子,其产生可能有关。
针对脂质纳米粒子(LNPs)的脂质成分,特别是阳离子脂质。因此,我们推测在我们的研究中,白介素-6(IL-6)的诱导可能促进树突状细胞(GC)的反应,这进一步有助于由狂犬病mRNA疫苗接种提供的持续体液免疫反应和长期保护。需要更多的研究和证据来阐明IL-6在这一过程中的确切作用。
研究表明,狂犬病疫苗依赖于T细胞来塑造免疫反应。CD8+ T细胞在消除被病毒感染的细胞和细胞内病毒中起着关键作用,而CD4+ T细胞则协助B细胞成熟并促进CD8+ T细胞的功能。我们的结果显示,mRNA疫苗在小鼠和恒河猴中诱导出比灭活疫苗更高的特异性IFN-γ+细胞免疫反应。同时,我们发现,在灭活疫苗组中,即使没有刺激,也存在激活的T细胞(见图3),这些细胞对抗原再刺激的反应不佳,这可能不利于将来对抗病原体的特定保护。需要研究来调查这些细胞是否是非特异性地被激活,或者在接种疫苗后未能有效地转变为记忆表型。
核苷酸修饰是mRNA治疗临床转化的一个里程碑。在这里,我们比较了含有修饰或未修饰核苷酸的mRNA疫苗。我们发现,m1up修饰提高了G蛋白在体外的表达效率和半衰期。在小鼠模型中,m1up修饰赋予了更高的抗体滴度,但未观察到显著差异。除了分泌IFN-γ的T细胞频率外,修饰和未修饰mRNA接种组之间的B细胞和T细胞反应几乎相当。如先前报道,核苷酸修饰并未显示出相对于未修饰狂犬病mRNA在小鼠中的明显优势。由于动物来源有限,我们未在非人灵长类同种动物中进行比较。最近的一项研究表明,m1μ修饰诱导了核糖体移码并影响了翻译准确性。需要进一步研究m1up掺入狂犬病mRNA疫苗的必要性,应在非人灵长类模型中进行。
总之,我们的数据表明,狂犬病mRNA疫苗在小白鼠和恒河猴中的表现均优于已批准的灭活疫苗。这突显了mRNA平台在开发下一代狂犬病疫苗方面的潜力。
材料与方法
细胞、病毒和动物
HEK293T和BHK-21细胞在37°C下、5%二氧化碳细胞培养箱中用含有10%胎牛血清(VivaCell,中国)和1%青霉素和链霉素(NCM Biotech,中国)的DMEM培养基(VivaCell,中国)中培养。用于挑战的CVS-11狂犬病病毒株和街毒狂犬病病毒株HuNPB3由中国农业科学院长春兽医研究所保存。从中国北京维通利华实验动物技术有限公司购买的雌性BALB/c小鼠(6-8周龄),在实验前接受了为期一周的预适应。恒河猴(5只雌性,4只雄性)被安置在中国药科大学新药安全评价中心的动物设施中。实验前,它们根据体重和性别随机分组(mRNA组:2只雄性和2只雌性;灭活疫苗组:1只雄性和2只雌性;磷酸盐缓冲液组:1只雄性和1只雌性)。所有动物实验均依据中国药科大学的伦理指南执行(批准编号l2023013)。
mRNA和脂质纳米粒(LNP)制备
狂犬病mRNA疫苗基于密码子优化的CTN-1株G蛋白序列(GenBank:FJ959397.1)构建。抗原序列由中国基因生物技术有限公司合成,包括启动子区域和UTR。线性化DNA片段通过PCR从模板扩增,然后由T7 RNA聚合酶(美国马萨诸塞州纽伯里公司)体外转录成mRNA。在转录过程中加入正常尿苷或m1up(美国加利福尼亚州特力联公司)。随后,通过RNA纯化试剂盒(纽伯里公司)纯化mRNA。经琼脂糖凝胶鉴定后,将mRNA储存在-80°C下。
通过微流体方法将狂犬病mRNA包封在脂质纳米粒中。简要来说,将mRNA稀释在柠檬酸盐缓冲液(pH 4.0)中,然后将可离子化脂质、二棕榈酰磷脂酰胆碱、胆固醇和聚乙二醇2000-二甲基丙烯酸酯(PEG2000-DMG)按摩尔比50/10/38.5/1.5溶解于乙醇中制成脂质混合物。接着,通过微流体芯片(中国苏州文皓微流体技术有限公司)以3:1的流速比将mRNA溶液与脂质混合物溶液混合。经过使用50 kD超滤管(德国Sartorius公司)进行离心过滤后,收集LNP-mRNA混合物,并使用Quant-iT RiboGreen RNA检测试剂盒(美国马萨诸塞州Thermo Scientific公司)分析包封效率。颗粒大小通过动态光散射(DLS)检测。
免疫策略
由国内疫苗生产商慷慨捐赠的纯化Vero细胞培养疫苗(一种用巴斯德病毒(PV)株制备的人类用狂犬病疫苗,冻干,标记效力为1剂量≥2.5IU,无佐剂)作为阳性对照。为简便起见,稿件中将此疫苗称为“灭活”疫苗。小鼠以0.5、1.5或5微克狂犬病mRNA肌内(im.)注射两次,间隔3周。在指示日期纵向采集血清样本进行抗体滴定(图2A)。在指示的时间点处安乐死小鼠进行相应的分析。接受相同免疫程序的灭活疫苗(0.25IU)或PBS(100微升)的小鼠分别作为阳性和阴性对照。
对于恒河猴实验,在第0天和第28天肌内(i.m.)注射一次狂犬病mRNA疫苗(30微克)、灭活疫苗(2.5IU)或PBS。如图5A所示进行后续检查。
酶联免疫吸附试验(ELISA)
每孔加入100纳克/孔的CTN-1-G蛋白(由中国南京德泰生物工程公司合成),在96孔板(美国纽约康宁公司)中过夜4°C孵育后,然后用含3%牛血清白蛋白(BSA,中国索莱宝公司)的PBS在37°C封闭ELISA板2小时。洗涤后,包被的平板依次与2倍系列稀释的小鼠血清(对于IgG1、IgG2a、IgG2b和IgG3抗体分析,血清稀释为1:8000)在37°C下孵育2小时。五次洗涤后,加入辣根过氧化物酶(HRP)偶联的抗小鼠IgG、IgG2a、IgG2b、IgG3抗体(美国阿拉巴马州南特生物科技公司)、HRP偶联的抗小鼠IgG1抗体(中国武汉三鹰生物技术有限公司)或HRP偶联的抗猴IgG(美国马萨诸塞州剑桥抗体公司),并在37°C下孵育1小时。然后加入TMB底物(纽康比生物科技公司),室温下孵育10-15分钟。通过加入终止缓冲液(纽康比生物科技公司)终止反应,并立即在450纳米处读取吸光度以计算滴定终点。
病毒中和试验
荧光抗体病毒中和(FAVN)用于检测血清中的中和抗体滴度。简而言之,将灭活血清样本(每孔50微升)按3倍系列稀释从第1行加到第6行的一个96孔板中。同时加入标准血清(0.5IU/毫升)作为参考。接着,向每个孔中加入100个荧光焦点单位(FFU)的CVS-11狂犬病病毒,然后在5%二氧化碳培养箱中37°C孵育1小时。随后,向每个孔中加入2万个BHK-21细胞,继续孵育48小时,然后用冰冷的80%丙酮处理30分钟。用FITC标记的抗狂犬病病毒N蛋白抗体染色后,通过荧光显微镜观察结果,并与参考血清进行比较计算。
病毒攻击
用狂犬病mRNA疫苗免疫雌性BALB/c小鼠(6-8周龄),接种剂量为0.5、1.5微克)或灭活疫苗(0.25IU)。接受PBS注射的小鼠被用作阴性对照。免疫后14天,小鼠通过腹腔注射街毒RABV株HuNPB3(100倍最小致死剂量50),随后观察动物的存活情况和体重。
样本采集和处理
小鼠在处死后浸入75%乙醇中5分钟。收集脾脏并通过密度梯度离心法处理以获得单细胞悬浮液,使用淋巴细胞分离介质(中国达科为),按照制造商的说明操作。为了从恒河猴中获得外周血单个核细胞(PBMC),将外周血用PBS按1:1稀释,然后轻轻铺在人类淋巴细胞分离介质上。密度梯度离心后,从白色膜层收集PBMC。
流式细胞术
流式细胞术按先前报道的方法进行。在染色前,单细胞悬浮液用抗CD16/CD32单克隆抗体(美国BD生物科学公司)或5%牛血清白蛋白(BSA)封闭。数据使用美国BD生物科学公司的BD FACSCelesta流式细胞仪收集,并使用FlowJo 10(美国树星公司)进行分析。
AIM+T细胞
用CTN-1-G蛋白(10微克/毫升)刺激小鼠脾细胞20小时,然后在室温避光条件下用可固定活力染料染色15分钟。洗涤后,细胞在4°C下与抗体鸡尾酒共孵育30分钟。该组合在表S1中提供。
免疫染色法
首先用或不用CTN-1-G蛋白(10微克/毫升)刺激小鼠的脾细胞或恒河猴的外周血单个核细胞16小时,然后在GolgiStop(BD生物科学公司)存在的情况下继续培养4小时。使用固定/渗透性溶液试剂盒(BD生物科学公司)按照制造商的说明进行表面和细胞内染色。该组可在表S2和S3中获得。
浆母细胞和生发中心B细胞
为了制备CTN-1-G探针,将生物素化的CTN-1-G蛋白与APC-、PE-或BV421-链霉亲和素以4:1的摩尔比偶联。首先在室温避光条件下用可固定活性染料染色小鼠的脾细胞或恒河猴的外周血单个核细胞15分钟,然后用CTN-1-G探针孵育30分钟。洗涤后,细胞与抗体鸡尾酒孵育30分钟。该组可在表S4-S6中获得。
酶联免疫斑点试验
每孔加入0.4百万个鼠脾细胞,用或不用CTN-1-G蛋白(5微克/毫升)孵育20小时。使用商业试剂盒(瑞典Mabtech公司)按照制造商的说明评估分泌IFN-γ或IL-4的T细胞数量。使用BCIP/NBT底物显色斑点,并用CTL免疫斑点S6分析仪计数。
血浆细胞因子的定量
使用美国乔治亚州的RayBiotech公司的细胞因子阵列来定量恒河猴血清中的细胞因子。简而言之,向芯片的每个孔中加入100微升样品稀释液,然后在室温下孵育1小时。随后,我们移除了加入的100微升标准溶液和两倍稀释的血清样本的缓冲液,然后在4摄氏度下过夜孵育。洗涤玻片后,每孔加入80微升检测抗体工作液,然后在室温下孵育75分钟。再次洗涤玻片后,每孔加入80微升Cy3-链霉亲和素,并在室温下避光孵育1小时。使用动态激光共聚焦扫描仪(Innopsys,伊利诺伊州,美国)检测荧光,并根据标准曲线计算细胞因子浓度。
统计分析
所有统计分析均使用GraphPad Prism(v.8.0,加利福尼亚州,美国)进行。p值小于0.05被认为具有统计学显著性(p<0.05,p<0.01,p<0.001,p<0.001;ns,无显著性)。
数据和代码可用性
所有数据均包含在论文或补充信息中。额外数据可根据合理请求向相应作者索取。
致谢
本工作得到了国家自然科学基金(82104219、8227131745和3247080161)、江苏省自然科学基金(BK20221521)、中央高校基本科研业务费(2632022ZD04)、中国药科大学天然药物国家重点实验室项目(SKLNMZZ202309)和中国国家重点研发计划(2024YFA0916700和2024YFA0918600)的支持。作者感谢中国药科大学基础医学院与临床药学院新药安全评价研究中心及核心设施在本研究中的仪器和技术支持。
作者贡献
XT, Y.Y, AL, F.Y, LZ, 和 W.Z 设计并监督了本研究,并修订了稿件。Y.W., S.W., LH., W.M., F.L., XZ, Y.Z, D.Y.Y.H., 李依丹, LR., 和 李颖参与了实验的开展和数据分析和处理。XT. 和 Y.W. 撰写了稿件。所有作者阅读并批准了提交的稿件。
利益声明
作者没有需要声明利益冲突的情况。
生成式AI和AI辅助技术在写作过程中的应用声明
在准备本工作的过程中,作者使用了ChatGPT进行英文校对。在使用该工具后,作者根据需要对内容进行了审查和编辑,并对出版物的内容承担全部责任。
补充信息
补充信息可在线获取:https://doi.org/10.1016/jymthe.202412.041。
参考文献
https://pubmed.ncbi.nlm.nih.gov/39741409/
Hits: 148
- 狂犬病(2025年6月更新)
- 狂犬病 你应该知道的
- 预防和治疗狂犬病的狂犬病疫苗
- 狂犬病可能成为历史
- 看看每种疫苗:狂犬病疫苗
- 阿拉巴马州妇女在从车上卸下杂货时被狂犬病狐狸咬伤:“公共卫生威胁”
- 应对狂犬病
- 全球疫苗免疫联盟将促进全球疫苗免疫联盟、世卫组织和阿联酋50多个国家获得拯救生命的人类狂犬病疫苗
- 美国CDC发布了将狗带入美国的新规定,旨在防止狂犬病
- 全国狂犬病监测方案
- 2024年东帝汶首例狂犬病确诊病例
- WHO实况报道 狂犬病
- 狂犬病
- 狂犬病疫苗接种:给医疗保健提供者的信息
- 特定感染的流行病学和控制:狂犬病
- 狂犬病疫苗接种的注意事项或禁忌症
- ACIP 使用减少(4剂)疫苗计划进行暴露后预防以预防人类狂犬病
- 实施良好的狂犬病暴露后预防方案
- ACIP 使用修改后的暴露前预防疫苗接种计划预防人类狂犬病,美国,2022年
- 西班牙犬类狂犬病免疫保护的影响因素
- 减少犬的进口等待期-对狂犬病恐惧科学合理的变化
- 老年人对狂犬病暴露后预防无应答:挑战和方法
- 狂犬病疫苗的效力、免疫原性和有效性:体外和体内方法
- 狂犬病人免疫球蛋白 (HRIG)
- “震惊”的妇女,超过6,000美元的救命狂犬病治疗账单
- 负鼠感染狂犬病毒对这种病毒在城市环境中的传播敲响了警钟
- 索马里蜜獾感染未报告的首例疑似人狂犬病病例研究
- 两名狂犬病研究人员解释如何保护自己免受这种疾病的侵害
- 临床医生常见的狂犬病问题
- 狂犬病疫苗
- 世界卫生组织关于狂犬病的常见问题
- 狂犬病实况报道
- 狂犬病
- 狂犬病是一种古老的、不可预测的、可能致命的疾病——两位狂犬病研究人员解释了如何保护自己
- 狂犬病疫苗短缺:给卫生专业人员的建议
- 7例被狂犬咬伤者接种狂犬病疫苗的效果观察
- 全球狂犬病风险:狂犬病疫苗短缺期间的实用建议
- 巴西一个土著村庄被蝙蝠掠夺的高风险:警告狂犬病在马卡利人中重新出现
- 评估在澳大利亚蝙蝠狂犬病病毒和狂犬病暴露后预防中血清学的益处
- 蝙蝠处理者的狂犬病暴露前预防
- 基于狂犬病中和抗体水平的狂犬病加强剂时机指南
- 施用狂犬病疫苗和免疫球蛋白
- 为狂犬病暴露后预防开发的单克隆抗体鸡尾酒疗法
- 狂犬病预防处置常见问题解答(成大生物)
- 世界卫生组织狂犬病专家磋商会 第三版报告
- 世卫组织对狂犬病的看法
- 新型狂犬病疫苗
- 狂犬病病毒可以抗强大的天然免疫
- 狂犬病的被动免疫
- 亚洲狂犬病预防
- 秘鲁高风险人群的大规模狂犬病暴露前预防
- 人用狂犬病疫苗
- 野生动物狂犬病疫苗
- 狂犬病和狂犬病疫苗的历史
- 狂犬病的生命周期、传播和发病机制
- 狂犬病暴露后管理指南(英国 2023年1月)
- 2022年印度马哈拉施特拉邦一名幼儿出现临床狂犬病症状后存活
- 第27章:狂犬病
- 狂犬病:问题和答案
- 《狂犬病暴露预防处置工作规范(2023年版)》解读问答
- 狂犬病暴露预防处置工作规范(2023年版)
- 狂犬病
- 狂犬病相关问题解答
- 疾病控制中心加强了对进口狗的狂犬病管理,原因如下
- 关于狂犬病
- 一名加州艺术教师在教室遭遇蝙蝠后死于狂犬病
- WHO 实况报道 动物咬伤 2024年1月
- 疫苗接种和免疫 狂犬病
- 世卫组织推荐皮内途径用于狂犬病暴露后预防
- 狂犬病疫苗:世卫组织立场文件–2018年4月
- WHO实况报道 狂犬病 2024年6月
- 狂犬病-结束忽视, 实现可持续发展目标 被忽视的热带病路线图 2021-2030
- 动物狂犬病
- 宠物主人对疫苗的犹豫越来越多——一位公共卫生专家解释了为什么这很重要
- 兽用狂犬病疫苗
- 何时寻求狂犬病医疗护理
- 猫
- 关于获得人类狂犬病疫苗的社会正义观点
- 评估新的狂犬病暴露后预防(PEP)方案或疫苗
- 卫生和福利部-狂犬病
- 狂犬病暴露后预防用单克隆抗体开发的最新进展:临床开发管线现状综述
- 世卫组织对狂犬病免疫接种的立场 – 2018 年最新情况
- 恐水症:1874 年的狂犬病和改变一切的救命疫苗
- 新的人类狂犬病疫苗正在研发中
- 为什么消除狂犬病?
- 为什么现在要根除狂犬病?
- 狂犬病-NFID
- 为狗接种疫苗 – 它们不会因接种疫苗而患上自闭症
- 宠物主人版 狗的狂犬病
- 猫的狂犬病
- 狂犬病-国际脑炎协会
- 器官移植后如何感染狂犬病?
- 器官移植患者感染狂犬病后死亡
- 人类狂犬病疾病负担:Rabipur®疫苗的潜在预防作用
- 印度东北部农村的狂犬病:强调同一健康方法紧迫性的病例报告
- 狂犬病毒糖蛋白的深度突变扫描确定了突变限制和抗体逃逸突变
- 人类狂犬病:一种治疗方法
- 通过受感染狗唾液污染的隐形眼镜传播狂犬病病毒(病例报告)
- 越南暴露前预防计划中基于灭活 vero 细胞的狂犬病疫苗 (Rabivax-S) 的安全性和免疫原性
- 狂犬病生物制剂在 III 类动物暴露儿科患者中的安全性和临床疗效的前瞻性研究
- 重复狂犬病疫苗接种揭示人类中和抗体的分子趋同
- 狂犬病病毒的感染和预防
- 关于 Zagreb 或 Essen 方案后加强疫苗接种的免疫原性和安全性的 3 期临床试验
- 通过 RABV-G 结构域筛选和异源初免-加强免疫优化狂犬病 mRNA 疫苗功效
- 狂犬病实验室诊断的最新情况
- 三色蝠相关狂犬病毒引起的一例人狂犬病的报道
- 一种核苷修饰的狂犬病mRNA疫苗在小鼠和非人灵长类动物中诱导出持久而全面的免疫反应
- 狂犬病的识别和治疗:病例报告和讨论
- 人类狂犬病的诊断困难:病例报告和文献回顾
- 狂犬病感染的边缘系统症状
- 狂犬病脑炎的生存率
- 人类狂犬病:2016年更新
- 突尼斯一名狂犬病幸存儿童:病例报告
- 肯尼亚蝙蝠携带的Duvenhage病毒导致人类狂犬病死亡:昏迷诱导、氯胺酮和抗病毒药物治疗无效
- 病例报告:狂犬病幸存者长期随访中的神经功能恢复与血清学反应
- 蝙蝠和食肉动物中狂犬病的比较病理学及其对人类溢出影响
- 病例报告:狂犬病生存者:印度病例系列
- 评估人类抗狂犬病疫苗的反应:一种快速、高通量适应性、基于假病毒的中和试验,作为快速荧光病灶抑制试验 (RFFIT) 的替代方案
- 病例报告:狂犬病幸存者长期随访中的神经功能恢复与血清学反应
- 巴西巴伊亚州狂犬病爆发的流行病学与分子分析
- 俄罗斯下诺夫哥罗德地区动物狂犬病(2012-2024年):描述性与预测性流行病学分析
- 巴西东南部一只黑鼠耳蝠(黑鼠耳蝠属物种Myotis nigricans)感染狂犬病病毒、科勒巴尔通体菌以及婴儿利什曼原虫的混合感染情况
- 2013-2023 年悉尼大都会居民狂犬病暴露后治疗:回顾性病例系列分析
- 用肌肉靶向脂质纳米颗粒配制的狂犬病病毒 G 蛋白 mRNA 在小鼠体内的免疫原性
- 关于狂犬病的 One Health Lens:马拉维利隆圭农村狂犬病病毒的人类-蝙蝠相互作用和基因组见解
- 阿根廷低地貘 (Tapirus terrestris) 麻痹狂犬病的首次报告
- 由表达狂犬病糖蛋白的非复制型病毒载体痘苗病毒天坛株改良株(MVA)和腺病毒5型(Ad5)所诱导产生的对狂犬病的保护作用
- 印度野生猫科动物中狂犬病毒的免疫病理与分子筛查:揭示从野外到城市的溢出效应
- 韩国貉 (Nyctereutes procynoide koreensis) 接种了含有 ERAGS-GFP 毒株的新型狂犬病诱饵疫苗的保护性抗体反应
- 宠物护理 疫苗
- 为您的宠物接种疫苗
- 狂犬病感染:如何保护您的孩子
- 新型狂犬病暴露后预防单克隆抗体鸡尾酒疗法的安全性和耐受性
- “愤怒的狂犬病”人类图画案例
- 蝙蝠咬伤
- 研究蝙蝠病毒如何帮助预防人畜共患疾病
- 狂犬病:症状繁多、宿主广泛的恶魔病毒——罗德尼·罗德解读
- 狂犬病的同一健康:不仅适用于动物
- 狂犬病和蛔虫:浣熊相关传染病
- 丽莎病毒 很罕见,但致命。如果蝙蝠咬你怎么办?
- 海地在四个省开展新的疫苗接种运动,加强防治犬类狂犬病
- 家养及其他动物狂犬病病毒感染的挑战:病毒学层面的困惑、惊愕、懊恼与好奇交织
- 双重战斗:狂犬病和蛇咬伤在东南亚的毒害
- 狂犬病在城市无人管理的猫群中爆发 — 马里兰州,2024 年 8 月
- 尼泊尔正力争在2030年前消灭狂犬病,但领导人表示,现在是加大疫苗投资的时候了。
- 现代狂犬病疫苗的诞生历程
- 一种基于mRNA的新型狂犬病病毒多表位疫苗:通过反向疫苗学和免疫信息学进行计算机设计
- 快速荧光灶抑制试验(RFFIT)的验证与评价 —— 作为狂犬病疫苗相对效价间接检测方法的中和抗体定量检测:NIH 试验的替代方案
- 暴露后狂犬病预防失败案例研究
- 2021年明尼苏达州一起经确认的人畜共患病暴露后,疑似因宿主介导导致暴露后预防失败的致命人类狂犬病感染病例
- 给野生动物投喂疫苗:口服狂犬病疫苗的故事
- 降低肯尼亚西部臭名昭著的狗市狂犬病风险
- 印度的狂犬病
- 海地的狂犬病
- 2024年10月至2025年2月,美国发生通过实体器官移植传播狂犬病的病例,该器官移植供体未确诊患有狂犬病
- 两例死亡与狂犬病感染的肾移植有关
- 明尼苏达州一家奶牛场的牛群中爆发狂犬病聚集性疫情——2024年
- 摩洛哥的狂犬病
- 2024年肯塔基州和俄亥俄州输入性人类狂犬病病例
- 2024年明尼苏达州和加利福尼亚州人类狂犬病死亡人数
- 反疫苗人士对宠物狂犬病 RNA 疫苗的虚假宣传
- 新型口服狂犬疫苗能否帮助我们最终终结这种疾病?
- NACI 2026 年 1 月 30 日发布的声明摘要:狂犬疫苗用于暴露前预防 (PrEP) 的最新指南
- 狂犬疫苗的给药途径:数据比较
- 猫会得狂犬病吗?
- 2022-2024年中欧狂犬病疫情再度出现
- 研讨会报告:改进狂犬病血清学方法和程序的标准化
- 我们来谈谈狂犬病检测
- 实体器官移植受者狂犬病:近期病例报告的启示
- 潜在狂犬病动物暴露评估最新指导意见
- 小罗伯特·F·肯尼迪想让狂犬病再次伟大
- 狂犬病疫苗研发的新篇章
- NACI 2026 年 4 月声明摘要:关于评估接触潜在狂犬病动物的最新指南
- 接种过疫苗的狗还会感染狂犬病吗?
- 仓鼠会得狂犬病吗?
- 如果已经接种过狂犬病疫苗,为什么还需要接种加强针?