
Contents
- 1 数据库
- 2 基于流感病毒
- 3 产品名
- 4 RSM2eFP孢子疫苗
- 5 产品名
- 6 普罗塔克
- 7 产品名
- 8 丙马流感
- 9 产品名
- 10 NAe-HA 和 M2e-HA
- 11 产品名
- 12 基于 HA 的马赛克整体 …
- 13 产品名
- 14 ΔNS1病毒
- 15 产品名
- 16 LAIV+4M2e
- 17 产品名
- 18 Reassortant LAIV 带 …
- 19 产品名
- 20 H1、H3 眼镜蛇 IIV
- 21 产品名
- 22 α-1,3-GT
- 23 产品名
- 24 S-流感
- 25 产品名
- 26 TX98-129型
- 27 产品名
- 28 ctLAIV的
- 29 产品名
- 30 科达瓦克斯
- 31 产品名
- 32 嵌合 HA 结构
- 33 产品名
- 34 BPL-1357型
- 35 产品名
- 36 M2SR系列
- 37 产品名
- 38 德尔塔FLU
- 39 基于核酸
- 40 产品名
- 41 ta-RNA、sa-RNA
- 42 产品名
- 43 优化的 M2e DNA …
- 44 产品名
- 45 cGAMP佐剂 …
- 46 产品名
- 47 NP mRNA的
- 48 产品名
- 49 四价 HA mRNA
- 50 产品名
- 51 SAM-GM-CSF + SAM-NP
- 52 产品名
- 53 pABOL配制的saRNA …
- 54 产品名
- 55 HA-F DNA疫苗
- 56 产品名
- 57 mRNA/LNP疫苗
- 58 产品名
- 59 H1c-mRNA-LNP
- 60 产品名
- 61 mRNA-流感
- 62 产品名
- 63 保守的T细胞 …
- 64 产品名
- 65 DNA 引物-亚基 …
- 66 产品名
- 67 MLN-mRNA的
- 68 产品名
- 69 PAPB1M1和NPPB2M2 …
- 70 产品名
- 71 pEx 4M2e
- 72 产品名
- 73 DNA 构建体 …
- 74 产品名
- 75 多价DNA疫苗
- 76 产品名
- 77 自扩增mRNA
- 78 产品名
- 79 多个 HA-DNA
- 80 产品名
- 81 mRNA-LNP模型
- 82 产品名
- 83 多抗原DNA …
- 84 产品名
- 85 sa-mRNA双反子疫苗
- 86 产品名
- 87 mRNA-1230 (英文)
- 88 产品名
- 89 DCVC H1 HA mRNA-LNP
- 90 产品名
- 91 mRNA-LNP H1ssF_3928 VRC
- 92 产品名
- 93 saRNA核糖核酸
- 94 产品名
- 95 微共识DNA疫苗
- 96 产品名
- 97 流感-SV-mRNA
- 98 产品名
- 99 mRNA-1011 和 mRNA-1012
- 100 产品名
- 101 mRNA-1020 和 mRNA-1030
- 102 产品名
- 103 基于modRNA的组合
- 104 产品名
- 105 mRNA构建体
- 106 产品名
- 107 mRNA-1010 (英语)
- 108 产品名
- 109 mRNA-1083 (英文)
- 110 产品名
- 111 改良的mRNA疫苗
- 112 非VLP纳米颗粒
- 113 产品名
- 114 M2E/CpG-ODN/TMC
- 115 产品名
- 116 HA三聚体
- 117 产品名
- 118 3M2e-rHF纳米颗粒
- 119 产品名
- 120 基于 M2e …
- 121 产品名
- 122 自组装蛋白 …
- 123 产品名
- 124 双层蛋白 …
- 125 产品名
- 126 HA/GP纳米颗粒
- 127 产品名
- 128 ISCOMs/MPLA 佐剂 …
- 129 产品名
- 130 脂质体肽 …
- 131 产品名
- 132 3M2e-T4纳米粒子
- 133 产品名
- 134 Mini-HA-LS纳米疫苗
- 135 产品名
- 136 佐剂 …
- 137 产品名
- 138 3MCD-f纳米疫苗
- 139 产品名
- 140 CDh-f纳米颗粒
- 141 产品名
- 142 HMN-PP型
- 143 产品名
- 144 双氟菌素
- 145 产品名
- 146 BP26-M2e纳米桶
- 147 产品名
- 148 PapMV-sM2e型 …
- 149 产品名
- 150 HA-脂质体
- 151 产品名
- 152 Helix-A阀杆 …
- 153 产品名
- 154 组合的 …
- 155 产品名
- 156 自组装 …
- 157 产品名
- 158 自组装 …
- 159 产品名
- 160 PLGA纳米颗粒
- 161 产品名
- 162 多价DNA …
- 163 产品名
- 164 M2e纳米团簇
- 165 产品名
- 166 结合疫苗和 …
- 167 产品名
- 168 CTA1-3M2e-DD (FPM2e)
- 169 产品名
- 170 IAV-纳米疫苗
- 171 产品名
- 172 Ace-DEX微粒
- 173 产品名
- 174 嵌合 M2e …
- 175 产品名
- 176 合成 …
- 177 产品名
- 178 鼻内NLP:NP
- 179 产品名
- 180 pH 响应性 NP …
- 181 产品名
- 182 AH3–GFP系列
- 183 产品名
- 184 VT-105型
- 185 产品名
- 186 EBS-UFV-001 (UFluA)
- 187 产品名
- 188 FluMos自组装纳米颗粒
- 189 产品名
- 190 HA2 HA-铁蛋白纳米颗粒
- 191 产品名
- 192 稳定的无头HA茎纳米颗粒
- 193 产品名
- 194 CIC疫苗
- 195 产品名
- 196 OVX836系列
- 197 产品名
- 198 纳米流感 (qNIV)
- 199 重组蛋白
- 200 产品名
- 201 产品名
- 202 产品名
- 203 NMHC系列
- 204 产品名
- 205 rM2e-ΔPly
- 206 产品名
- 207 M2e-H3 秸秆
- 208 产品名
- 209 rNA抗原
- 210 产品名
- 211 rNP 加 BPPcysMPEG
- 212 产品名
- 213 马赛克和嵌合 HA
- 214 产品名
- 215 核糖核酸
- 216 产品名
- 217 3M2e-3HA2-NP嵌合体 …
- 218 产品名
- 219 nM2Pr
- 220 产品名
- 221 LHNVD-105型
- 222 产品名
- 223 糖脂肽
- 224 产品名
- 225 HA茎蛋白
- 226 产品名
- 227 Clec9A-M2e型
- 228 产品名
- 229 3M2e-HA2-NP嵌合体 …
- 230 产品名
- 231 基于M2e的重组体 …
- 232 产品名
- 233 TAT-NP型
- 234 产品名
- 235 HAsd 蛋白
- 236 产品名
- 237 EFn-3xM2e-HA2+PA型
- 238 产品名
- 239 HA-T-AGM 蛋白
- 240 产品名
- 241 rNP+SLA-SE系列
- 242 产品名
- 243 α-DEC-205: M2e …
- 244 产品名
- 245 N1-I COBRA NA抗原
- 246 产品名
- 247 基于 H1 的 cHA
- 248 产品名
- 249 HA 迷你茎
- 250 产品名
- 251 单ADCC激活 …
- 252 产品名
- 253 三聚体HA-Fc
- 254 产品名
- 255 共轭 HA 茎
- 256 产品名
- 257 眼镜蛇 H1 HA
- 258 产品名
- 259 HA-clamp 蛋白
- 260 产品名
- 261 嵌合HA蛋白
- 262 产品名
- 263 加扰 HA (scrHA)
- 264 产品名
- 265 CapM2e+Advax-SM系列
- 266 产品名
- 267 B60-茎-8071
- 268 产品名
- 269 去糖基化 …
- 270 产品名
- 271 流感-v
- 272 产品名
- 273 G1 毫哈
- 274 产品名
- 275 FP-01.1型
- 276 产品名
- 277 多聚体-001 (M-001)
- 278 病毒样颗粒 (VLP)
- 279 产品名
- 280 NA2 VLP系列
- 281 产品名
- 282 HBc VLP
- 283 产品名
- 284 杂交融合蛋白 …
- 285 产品名
- 286 倒置 HA VLP
- 287 产品名
- 288 HA-VLP-细胞
- 289 产品名
- 290 HA/M1 VLP型
- 291 产品名
- 292 m-cNA-M2e VLP系列
- 293 产品名
- 294 M2e5x VLP(多个 …
- 295 产品名
- 296 N1 VLP系列
- 297 产品名
- 298 Cap-M2e VLP
- 299 产品名
- 300 T4-M2E VLP系列
- 301 产品名
- 302 P22-HA型头VLP(VLP)
- 303 产品名
- 304 坦迪氟1 VLP
- 305 产品名
- 306 cVLP的
- 307 产品名
- 308 NA-VLP系列
- 309 产品名
- 310 NA VLP
- 311 产品名
- 312 AP-M2e/三柄 VLP
- 313 产品名
- 314 M2e VLP MP
- 315 产品名
- 316 VLP鸡尾酒
- 317 产品名
- 318 嵌合细胞因子 …
- 319 产品名
- 320 马赛克 VLP
- 321 产品名
- 322 SpyTagged noro-VLP
- 323 产品名
- 324 基于VLP的疫苗
- 325 产品名
- 326 NvC-M2EX3型
- 327 产品名
- 328 中电HA茎
- 329 产品名
- 330 COBRA-VLP
- 331 产品名
- 332 HBc-4M2eh (Uniflu)
- 333 产品名
- 334 基于 M2e 的 VLP (ACAM-FluA)
- 335 产品名
- 336 四价 VLP (QVLP)
- 337 病毒载体
- 338 产品名
- 339 BWV-101型
- 340 产品名
- 341 RVJ-4M2eNP型
- 342 产品名
- 343 MVA载体疫苗
- 344 产品名
- 345 rAd/B-NP型
- 346 产品名
- 347 rMVA-k1-k2
- 348 产品名
- 349 A/NP+M2-rAd
- 350 产品名
- 351 ADC68-CR9114型
- 352 产品名
- 353 MVA-NP型
- 354 产品名
- 355 rAd-HA2型
- 356 产品名
- 357 AD-5-H1型
- 358 产品名
- 359 ChAdOx1 NP+M1 引物, …
- 360 产品名
- 361 rAd-SHA2
- 362 产品名
- 363 基于Ad的多表位
- 364 产品名
- 365 AAV 载体 HA 或 cHA
- 366 产品名
- 367 PIV5-不适用
- 368 产品名
- 369 惠氏/IL-15/5流感
- 370 产品名
- 371 MAV-1 哈
- 372 产品名
- 373 rVSV-EΔM-tM2e 和 …
- 374 产品名
- 375 H1 马赛克 HA
- 376 产品名
- 377 H2-康
- 378 产品名
- 379 HAdV-5-题词
- 380 产品名
- 381 ChAdOx1、NP+M1 和 MVA NP+M1 在异源初免-加强方案中的应用
- 382 产品名
- 383 AD4-H5-VTN型
- 384 产品名
- 385 VXA-A1.1口服片剂
- 386 产品名
- 387 NasoVAX系列
- 388 产品名
- 389 MVA-NP+M1 (VTP-100)
UNIVERSAL INFLUENZA VACCINE TECHNOLOGY LANDSCAPE
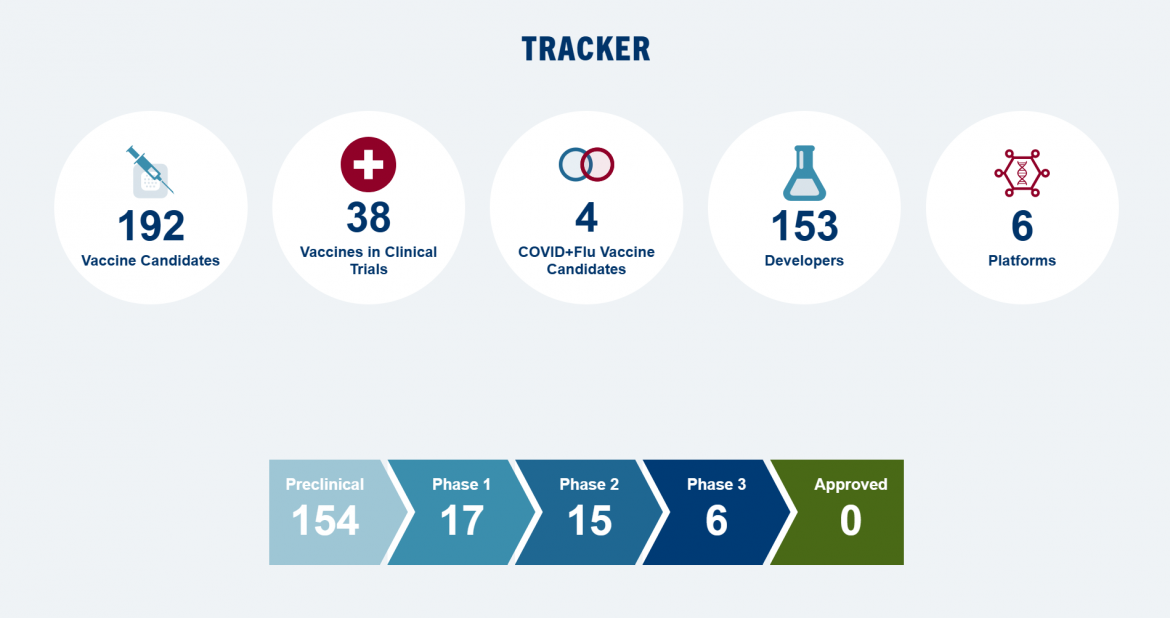
The Landscape是一个新型候选疫苗数据库,旨在与目前针对特定毒株的季节性流感疫苗相比,提供更广泛、更持久的保护,以抵御流行的和大流行性流感病毒。CIDRAP正在全球通用流感疫苗开发资助者联盟的支持下开发这一领域。
数据库最后更新于 2023 年 12 月 22 日
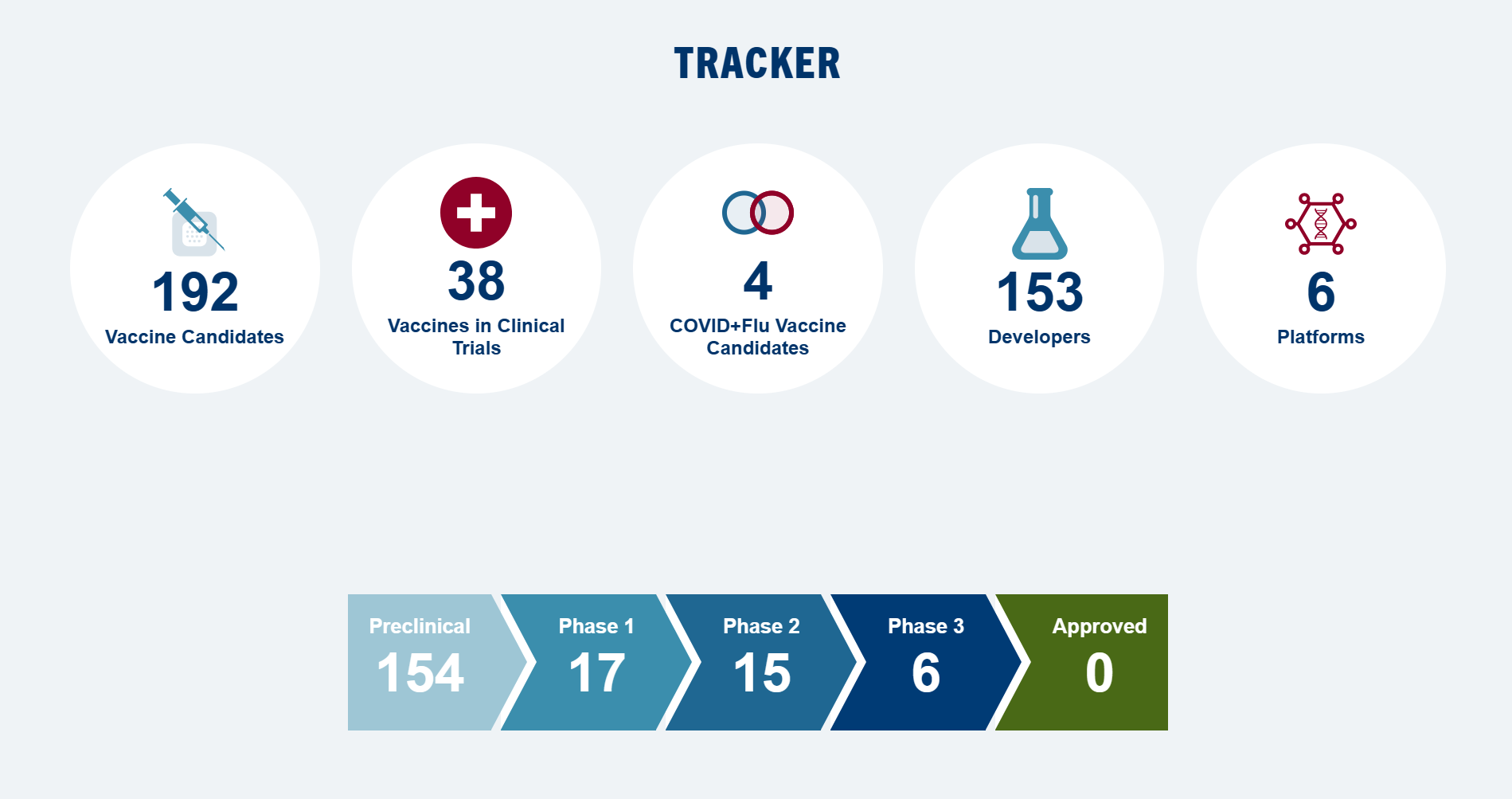
数据库
| 临床前 | 临床1期 | 临床2期 | 临床3期 | 批准 |
|---|---|---|---|---|
平台基于流感病毒 | ||||
| 13个疫苗 临床前 | 3个疫苗 临床1期 | 2个疫苗 临床2期 | 0个疫苗 临床3期 | 0个疫苗 批准 |
| 开发者 北京研究所 … 产品名RSM2eFP孢子疫苗开发者 中国科学院 … 产品名普罗塔克开发者 Gamma Vaccines (澳大利亚) 产品名丙马流感开发者 Georgia State University (US) 佐治亚州立大学 产品名NAe-HA 和 M2e-HA开发者 伊坎医学院 … 产品名基于 HA 的马赛克整体 …开发者 伊坎医学院 … 产品名ΔNS1病毒开发者 实验研究所 … 产品名LAIV+4M2e开发者 实验研究所 … 产品名Reassortant LAIV 带 …开发者 佐治亚大学(美国) 产品名H1、H3 眼镜蛇 IIV开发者 香港大学(香港 … 产品名α-1,3-GT开发者 牛津大学(英国) 产品名S-流感开发者 南达科他大学 … 产品名TX98-129型开发者 延世大学(南) … 产品名ctLAIV的 | 开发者 Codagenix (美国) 产品名科达瓦克斯状态 开发中 开发者 西奈山伊坎医学院(美国) 产品名嵌合 HA 结构状态 在开发 开发者 美国国家过敏和传染病研究所 … 产品名BPL-1357型状态 在开发 | 开发者 FluGen (美国) 产品名M2SR系列状态 在开发 开发者 Vivaldi Biosciences(美国) 产品名德尔塔FLU状态 在开发 | ||
平台基于核酸 | ||||
| 22个疫苗 临床前 | 6个疫苗 临床1期 | 5个疫苗 临床2期 | 3个疫苗 临床3期 | 0个疫苗 批准 |
| 开发者 BioNTech (德国) 产品名ta-RNA、sa-RNA开发者 中国科学院 … 产品名优化的 M2e DNA …开发者 Georgia State University (US) 佐治亚州立大学 产品名cGAMP佐剂 …开发者 根特大学(比利时) 产品名NP mRNA的开发者 Greenlight Biosciences (美国) 产品名四价 HA mRNA开发者 葛兰素史克(美国) 产品名SAM-GM-CSF + SAM-NP开发者 伦敦帝国理工学院 … 产品名pABOL配制的saRNA …开发者 吉林大学(中国) 产品名HA-F DNA疫苗开发者 Merck & Co. (美国) 产品名mRNA/LNP疫苗开发者 国家工程 … 产品名H1c-mRNA-LNP开发者 国家公共研究所 … 产品名mRNA-流感开发者 Saint Louis University (US) 圣路易斯大学 产品名保守的T细胞 …开发者 上海研究所 … 产品名DNA 引物-亚基 …开发者 上海研究所 … 产品名MLN-mRNA的开发者 上海市公共卫生 … 产品名PAPB1M1和NPPB2M2 …开发者 斯洛伐克科学院 … 产品名pEx 4M2e开发者 国家科学研究中心 … 产品名DNA 构建体 …开发者 史坦斯血清研究所 … 产品名多价DNA疫苗开发者 明尼苏达大学(美国) 产品名自扩增mRNA开发者 奥斯陆大学(挪威) 产品名多个 HA-DNA开发者 宾夕法尼亚大学 … 产品名mRNA-LNP模型开发者 University of Washington (US) 华盛顿大学 产品名多抗原DNA … | 开发者 CSL Seqirus (美国) Arcturus Therapeutics(美国) 产品名sa-mRNA双反子疫苗COVID + 流感 开发者 Moderna (美国) 产品名mRNA-1230 (英文)状态 在开发 开发者 美国国家过敏和传染病研究所 … 产品名DCVC H1 HA mRNA-LNP状态 在开发 开发者 美国国家过敏和传染病研究所 … 产品名mRNA-LNP H1ssF_3928 VRC状态 在开发 开发者 辉瑞(美国) 产品名saRNA核糖核酸状态 在开发 开发者 Wistar Institute (美国) 产品名微共识DNA疫苗状态 暂停 | 开发者 CureVac (德国) 葛兰素史克(美国) 产品名流感-SV-mRNA状态 在开发 开发者 Moderna (美国) 产品名mRNA-1011 和 mRNA-1012状态 在开发 开发者 Moderna (美国) 产品名mRNA-1020 和 mRNA-1030状态 在开发 COVID + 流感 开发者 辉瑞(美国) BioNTech (德国) 产品名基于modRNA的组合状态 在开发 开发者 赛诺菲巴斯德(美国) 产品名mRNA构建体状态 在开发 | 开发者 Moderna (美国) 产品名mRNA-1010 (英语)状态 在开发 COVID + 流感 开发者 Moderna (美国) 产品名mRNA-1083 (英文)状态 在开发 开发者 辉瑞(美国) BioNTech (德国) 产品名改良的mRNA疫苗状态 在开发 | |
平台非VLP纳米颗粒 | ||||
| 36个疫苗 临床前 | 4个疫苗 临床1期 | 2个疫苗 临床2期 | 1个疫苗 临床3期 | 0个疫苗 批准 |
| 开发者 农业研究 … 产品名M2E/CpG-ODN/TMC开发者 加州研究所 … 产品名HA三聚体开发者 中国科学院 … 产品名3M2e-rHF纳米颗粒开发者 复旦大学(中国) 产品名基于 M2e …开发者 佐治亚学院 … 产品名自组装蛋白 …开发者 Georgia State University (US) 佐治亚州立大学 产品名双层蛋白 …开发者 Georgia State University (US) 佐治亚州立大学 产品名HA/GP纳米颗粒开发者 Georgia State University (US) 佐治亚州立大学 产品名ISCOMs/MPLA 佐剂 …开发者 格里菲斯大学 … 产品名脂质体肽 …开发者 华中农业 … 产品名3M2e-T4纳米粒子开发者 华中农业 … 产品名Mini-HA-LS纳米疫苗开发者 印度科学研究所 … 产品名佐剂 …开发者 吉林大学(中国) 产品名3MCD-f纳米疫苗开发者 吉林大学(中国) 产品名CDh-f纳米颗粒开发者 吉林大学(中国) 产品名HMN-PP型开发者 KJ Biosciences (美国) 产品名双氟菌素开发者 韩国高等研究院 … 产品名BP26-M2e纳米桶开发者 拉瓦尔大学(加拿大) 产品名PapMV-sM2e型 …开发者 莫纳什大学(澳大利亚) 产品名HA-脂质体开发者 美国国家过敏研究所 … 产品名Helix-A阀杆 …开发者 国立台湾大学 … 产品名组合的 …开发者 俄罗斯科学院 … 产品名自组装 …开发者 斯克里普斯研究所 … 产品名自组装 …开发者 双威大学(马来西亚) 产品名PLGA纳米颗粒开发者 台州大学(中国) 产品名多价DNA …开发者 Texas Tech University (US) 德克萨斯理工大学 产品名M2e纳米团簇开发者 TRIA Bioscience(美国) 产品名结合疫苗和 …开发者 哥德堡大学 … 产品名CTA1-3M2e-DD (FPM2e)开发者 爱荷华大学(美国) 产品名IAV-纳米疫苗开发者 北卡罗来纳大学 … 产品名Ace-DEX微粒开发者 魁北克大学 … 产品名嵌合 M2e …开发者 魁北克大学 … 产品名合成 …开发者 罗切斯特大学 … 产品名鼻内NLP:NP开发者 范德比尔特大学(美国) 产品名pH 响应性 NP …开发者 Vaxsia Biomedical (台湾) 产品名AH3–GFP系列开发者 Versatope Therapeutics(美国) 产品名VT-105型 | 开发者 Emergent BioSolutions(美国) 产品名EBS-UFV-001 (UFluA)状态 在开发 开发者 美国国家过敏和传染病研究所 … 产品名FluMos自组装纳米颗粒状态 在开发 开发者 美国国家过敏和传染病研究所 … 产品名HA2 HA-铁蛋白纳米颗粒状态 在开发 开发者 美国国家过敏和传染病研究所 … 产品名稳定的无头HA茎纳米颗粒状态 在开发 | COVID + 流感 开发者 Novavax (美国) 产品名CIC疫苗状态 在开发 开发者 Osivax (法国) 产品名OVX836系列状态 在开发 | 开发者 Novavax (美国) Emergent BioSolutions(美国) 产品名纳米流感 (qNIV)状态 在开发 | |
平台重组蛋白 | ||||
| 36个疫苗 临床前 | 0个疫苗 临床1期 | 3个疫苗 临床2期 | 1个疫苗 临床3期 | 0个疫苗 批准 |
| 开发者 中央研究院(台湾) 产品名cHAmg 开发者 中国科学院 … 产品名M2e-CRM197 conjugates 开发者 中国科学院 … 产品名NMHC系列开发者 重庆医科大学 … 产品名rM2e-ΔPly开发者 Georgia State University (US) 佐治亚州立大学 产品名M2e-H3 秸秆开发者 根特大学(比利时) 产品名rNA抗原开发者 亥姆霍兹中心 … 产品名rNP 加 BPPcysMPEG开发者 伊坎医学院 … 产品名马赛克和嵌合 HA开发者 伊坎医学院 … 产品名核糖核酸开发者 韩国综合研究所 … 产品名3M2e-3HA2-NP嵌合体 …开发者 韩国综合研究所 … 产品名nM2Pr开发者 Longhorn 疫苗和 … 产品名LHNVD-105型开发者 莫纳什大学(澳大利亚) 产品名糖脂肽开发者 美国国家过敏研究所 … 产品名HA茎蛋白开发者 国立大学 … 产品名Clec9A-M2e型开发者 伊朗巴斯德研究所 … 产品名3M2e-HA2-NP嵌合体 …开发者 俄罗斯卫生部 … 产品名基于M2e的重组体 …开发者 上海研究所 … 产品名TAT-NP型开发者 西南交通大学 … 产品名HAsd 蛋白开发者 Texas Tech University (US) 德克萨斯理工大学 产品名EFn-3xM2e-HA2+PA型开发者 转化健康科学 … 产品名HA-T-AGM 蛋白开发者 特鲁多研究所(美国) 产品名rNP+SLA-SE系列开发者 Universidad Autónoma del 自治大学 … 产品名α-DEC-205: M2e …开发者 佐治亚大学(美国) 产品名N1-I COBRA NA抗原开发者 加纳大学(加纳) 产品名基于 H1 的 cHA开发者 香港大学(香港 … 产品名HA 迷你茎开发者 香港大学(香港 … 产品名单ADCC激活 …开发者 马里兰大学(美国) 产品名三聚体HA-Fc开发者 墨尔本大学 … 产品名共轭 HA 茎开发者 北卡罗来纳大学 … 产品名眼镜蛇 H1 HA开发者 昆士兰大学 … 产品名HA-clamp 蛋白开发者 罗切斯特大学 … 产品名嵌合HA蛋白开发者 威斯康星大学(美国) 产品名加扰 HA (scrHA)开发者 Vaxine (澳大利亚) 产品名CapM2e+Advax-SM系列开发者 厦门大学(中国) 产品名B60-茎-8071开发者 厦门大学(中国) 产品名去糖基化 … | 开发者 ConserV Bioscience(英国) Imutex (英国) 产品名流感-v状态 在开发 开发者 Janssen Vaccines and Prevention, J&J (荷兰) 产品名G1 毫哈状态 在开发 开发者 免疫靶向系统(Altimmune)(英国) 产品名FP-01.1型状态 暂停 | 开发者 BiondVax Pharmaceuticals (以色列) 产品名多聚体-001 (M-001)状态 暂停 | ||
平台病毒样颗粒 (VLP) | ||||
| 26个疫苗 临床前 | 2个疫苗 临床1期 | 0个疫苗 临床2期 | 1个疫苗 临床3期 | 0个疫苗 批准 |
| 开发者 Auburn University (美国) 产品名NA2 VLP系列开发者 中国科学院 … 产品名HBc VLP开发者 Emory University (美国) 产品名杂交融合蛋白 …开发者 佐治亚学院 … 产品名倒置 HA VLP开发者 Georgia State University (US) 佐治亚州立大学 产品名HA-VLP-细胞开发者 Georgia State University (US) 佐治亚州立大学 产品名HA/M1 VLP型开发者 Georgia State University (US) 佐治亚州立大学 产品名m-cNA-M2e VLP系列开发者 Georgia State University (US) 佐治亚州立大学 产品名M2e5x VLP(多个 …开发者 Georgia State University (US) 佐治亚州立大学 产品名N1 VLP系列开发者 河南省农业科学研究院 … 产品名Cap-M2e VLP开发者 华中农业 … 产品名T4-M2E VLP系列开发者 印第安纳大学(美国) 产品名P22-HA型头VLP(VLP)开发者 iQur (英国) 产品名坦迪氟1 VLP开发者 嘉兴大学(中国) 产品名cVLP的开发者 King Mongkut’s University of 蒙库特国王大学 … 产品名NA-VLP系列开发者 庆熙大学(韩国) 产品名NA VLP开发者 拉脱维亚生物医学研究 … 产品名AP-M2e/三柄 VLP开发者 Mercer University (US) 美世大学 产品名M2e VLP MP开发者 美国国家过敏研究所 … 产品名VLP鸡尾酒开发者 Nerome 研究所 … 产品名嵌合细胞因子 …开发者 中山大学(中国) 产品名马赛克 VLP开发者 坦佩雷大学(芬兰) 产品名SpyTagged noro-VLP开发者 TechnoVax(美国) 产品名基于VLP的疫苗开发者 马来西亚博特拉大学 … 产品名NvC-M2EX3型开发者 哥本哈根大学 … 产品名中电HA茎开发者 佐治亚大学(美国) 产品名COBRA-VLP | 开发者 俄罗斯科学院(俄罗斯) VA Pharma (俄罗斯) 产品名HBc-4M2eh (Uniflu)开发者 根特大学(比利时) 赛诺菲巴斯德(美国) 产品名基于 M2e 的 VLP (ACAM-FluA)状态 暂停 | 开发者 Medicago (加拿大) 产品名四价 VLP (QVLP)状态 暂停 | ||
平台病毒载体 | ||||
| 21个疫苗 临床前 | 2个疫苗 临床1期 | 3个疫苗 临床2期 | 0个疫苗 临床3期 | 0个疫苗 批准 |
| 开发者 Blue Water Biotech(美国) 产品名BWV-101型开发者 中国疾病预防控制中心(中国) 产品名RVJ-4M2eNP型开发者 Emergent BioSolutions(美国) 产品名MVA载体疫苗开发者 梨花女子大学(韩国) 产品名rAd/B-NP型开发者 联邦医学生物学 … 产品名rMVA-k1-k2开发者 美国食品和药物管理局 … 产品名A/NP+M2-rAd开发者 复旦大学(中国) 产品名ADC68-CR9114型开发者 德国感染中心 … 产品名MVA-NP型开发者 加拿大卫生部(加拿大) 产品名rAd-HA2型开发者 伊坎医学院 … 产品名AD-5-H1型开发者 伊坎医学院 … 产品名ChAdOx1 NP+M1 引物, …开发者 国立食品研究所 … 产品名rAd-SHA2开发者 Purdue University (US) 普渡大学 产品名基于Ad的多表位开发者 罗伯特·科赫研究所 … 产品名AAV 载体 HA 或 cHA开发者 佐治亚大学(美国) 产品名PIV5-不适用开发者 香港大学(香港 … 产品名惠氏/IL-15/5流感开发者 列日大学(比利时) 产品名MAV-1 哈开发者 曼尼托巴大学 … 产品名rVSV-EΔM-tM2e 和 …开发者 University of 大学 … 产品名H1 马赛克 HA开发者 University of 大学 … 产品名H2-康开发者 University of 大学 … 产品名HAdV-5-题词 | 开发者 牛津大学詹纳研究所(英国) 产品名ChAdOx1、NP+M1 和 MVA NP+M1 在异源初免-加强方案中的应用状态 暂停 开发者 美国国家过敏和传染病研究所 … 产品名AD4-H5-VTN型状态 暂停 | 开发者 Vaxart (美国) 产品名VXA-A1.1口服片剂状态 在开发 开发者 Altimmune (美国) 产品名NasoVAX系列状态 暂停 开发者 Vaccitech (英国) 产品名MVA-NP+M1 (VTP-100)状态 暂停 | ||
| 疫苗名称 | 开发商/地点 | 平台 | 阶段 | 方法和来源 |
|---|---|---|---|---|
| 3M2e-3HA2-NP嵌合蛋白 | 韩国生物科学和生物技术研究所(韩国) | 重组蛋白质 | 临床前的 | 靶向M2e、HA和NP病毒蛋白的嵌合蛋白;旨在诱导流感病毒特异性抗体应答、细胞毒性T淋巴细胞活性和抗体依赖性细胞毒性。 |
| 3M2e-HA2-NP嵌合亚单位 | 伊朗巴斯德研究所(伊朗) | 重组蛋白质 | 临床前的 | 含有来源于甲型流感病毒的M2e、HA和NP保守区的重组嵌合蛋白3M2e-HA2-NP。 马利基2023 (PMID: 37191741), 萨利赫2020 (PMID: 32152828), 法拉曼2019 (PMID: 30382564), Shokouhi 2018 (PMID: 29730305) |
| 3M2e-rHF纳米颗粒 | 中国科学院(中国) | 非VLP纳米粒子 | 临床前的 | 展示在铁蛋白纳米颗粒(重组人重链铁蛋白,rHF)上的M2e (3M2e)的串联拷贝;鼻内给药;旨在通过M2e特异性IgG抗体、T细胞免疫应答和粘膜分泌型IgA抗体刺激交叉保护性免疫。 |
| 3M2e-T4纳米颗粒 | 华中农业大学(中国) | 非VLP纳米粒子 | 临床前的 | 流感病毒M2e通过与Soc(小外壳蛋白)融合而插入噬菌体T4基因组中,产生重组噬菌体,并且在T4在大肠杆菌中繁殖期间,Soc-M2e蛋白自组装到噬菌体外壳上,形成3M2e-T4纳米颗粒。 |
| 3MCD-f纳米疫苗 | 吉林大学(中国) | 非VLP纳米粒子 | 临床前的 | 由两个保守表位(M2e和CDhelix)组成的无佐剂自组装流感纳米疫苗 |
| a/NP+M2-拉德 | 美国食品药品监督管理局(美国) | 病毒载体 | 临床前的 | 表达保守抗原NP和M2的复制缺陷型重组腺病毒5 (rAd)载体。 失误2023 (PMID: 37830821), 价格2022 (PMID: 35638848), Lo 2021 (PMID: 34226103), 达卡2021 (PMID: 33627390), 罗威尔2019 (PMID: 30986224), 罗威尔2018年奥运会(PMID: 29249542), 价格2018 (PMID: 30037481), Soboleski 2011 (PMID: 21789196), 价格2010 (PMID: 20976273) |
| AAV-矢量HA或cHA | 罗伯特·科赫研究所(德国) | 病毒载体 | 临床前的 | 表达流感病毒HA或嵌合HA的腺相关病毒(AAV)载体;旨在诱导广泛的保护性抗体。 |
| Ace-DEX微粒 | 北卡罗来纳大学(美国) | 非VLP纳米粒子 | 临床前的 | M2e-和环状GMP-AMP (cGAMP)包封的乙酰化葡聚糖(Ace-DEX)微粒(MP);抗原-佐剂组合旨在激活体液和细胞反应。 巴蒂2023 (PMID: 37458383), 巴蒂2023 (PMID: 36436743), 陈2018(:30261204) |
| ad-5 H1 | 马里兰大学西奈山伊坎医学院 | 病毒载体 | 临床前的 | 非复制型腺病毒(Ad)载体,编码分泌型H1 HA,旨在引发交叉反应性柄介导的细胞和体液免疫和保护。 |
| Ad4-H5-VTN | 美国国家过敏和传染病研究所 | 病毒载体 | 第一相 | 编码流感病毒H5 HA (Ad4-H5-Vtn)的复制型腺病毒4型以口服胶囊或通过扁桃体拭子或鼻腔喷雾给药,作为诱导针对流感糖蛋白的持久和系统性粘膜免疫的潜在平台。 松田隼2021 (PMID: 33529172), 亚历山大2012 (PMID: 22363572), 新闻稿2021年2月19日, NCT01443936, NCT01806909, 开发者网站:NIAID |
| AdC68-CR9114 | 复旦大学(中国) | 病毒载体 | 临床前的 | 表达CR9114的黑猩猩腺病毒载体AdC68,Cr 9114是从健康供体的外周血单核细胞中分离的广泛中和的单克隆抗体;CR9114针对不同类型的流感病毒。 |
| 佐剂化的纳米粒子融合构建体 | 印度科学研究所(印度) | 非VLP纳米粒子 | 临床前的 | 三聚体流感干结构域免疫原pH1HA10,展示在铁蛋白上,如蛋白质MsDps2、铁蛋白和囊素 卡尔2022 (PMID: 35720346), 安迪·纳亚尔·罗德里格斯2018年奥运会(PMID: 29863853) |
| AH3-GFP | Vaxsia Biomedical(台湾),马里兰大学巴尔的摩分校(美国) | 非VLP纳米粒子 | 临床前的 | 由AH3肽的功能增益突变体组成的自组装蛋白质纳米颗粒,其使得温度和盐依赖性蛋白质纳米颗粒热稳定,旨在刺激长期持续的体液免疫反应,该反应由在注射部位保持完整的热稳定蛋白质纳米颗粒介导 |
| AP-M2e/三茎VLP | 拉脱维亚生物医学研究中心(拉脱维亚) | 病毒样颗粒(VLP) | 临床前的 | 整合到噬菌体AP205 VLP平台中的基于HA三柄和三重M2e (3M2e)抗原的一组构建体;旨在针对季节性和新型流感病毒株提供广泛保护。 |
| B60-Stem-8071 | 厦门大学(中国) | 重组蛋白质 | 临床前的 | 一种移植有广泛中和抗体CR8071表位的HA干细胞疫苗,用于稳定干细胞HA结构域 |
| 双卤素 | KJ生物科学(美国),德克萨斯A&M大学(美国) | 非VLP纳米粒子 | 临床前的 | 在纳米颗粒蛋白上呈递两种保守抗原M2e和HA2的新型双结构域纳米颗粒融合蛋白;旨在诱导交叉保护性免疫反应。 |
| BP26-M2e纳米桶 | 韩国高等科学技术学院(韩国) | 非VLP纳米粒子 | 临床前的 | 带有旨在促进抗原特异性抗体产生的抗原的工程化单体BP26单体。 |
| BPL-1357 | 美国国家过敏和传染病研究所 | 基于流感病毒 | 第一相 | 鼻内或肌肉注射的全病毒疫苗,由四种非传染性、化学灭活的低致病性禽流感病毒株组成;旨在诱导粘膜免疫,类似于流感感染后的免疫反应,包括细胞和B细胞反应。 拉金2022 (PMID: 35943485), 公园2022, 公园2021, NCT05027932, 新闻稿2022年8月19日, 新闻稿2022年7月13日, 新闻稿2022年6月28日, 开发者网站:NIAID |
| BWV-101 | 蓝水生物技术公司(美国)、牛津大学(英国)、布坦坦研究所(巴西) | 病毒载体 | 临床前的 | 嵌合HA构建体在H1 HA头结构域中展示有限可变性的高免疫原性表位。 汤姆森2018 (PMID: 30242149), 新闻稿2023年4月24日, 新闻稿2022年8月24日, 新闻稿2022年5月19日, 新闻稿2021年9月22日, 新闻稿2019年1月31日, 专利, 开发者网站:蓝水生物科技 |
| Cap-M2e VLPs | 河南省农业科学院(中国)、郑州大学(中国) | 病毒样颗粒(VLP) | 临床前的 | 用Cap(PC v2的衣壳蛋白)VLPs展示M2e抗原;旨在针对甲型流感病毒株的不同亚型提供交叉保护。 |
| CapM2e+Advax-SM | 瓦欣语(澳大利亚) | 重组蛋白质 | 临床前的 | 用Advax-SM佐剂配制的基于M2e抗原的疫苗施用于母亲,旨在诱导对新生儿的被动母亲保护 |
| CDh-f纳米粒子 | 吉林大学(中国) | 非VLP纳米粒子 | 临床前的 | 来自HA茎的A螺旋(Ah)和CD螺旋(CDh)与铁蛋白单独或串联融合,产生Ah-f、CDh-f和(A + CD)h-f纳米颗粒(NPs),通过原核表达系统产生。 |
| cGAMP佐剂多价mRNA疫苗 | 佐治亚州立大学(美国) | 基于核酸的 | 临床前的 | 一种多价流感mRNA脂质纳米粒(LNP)疫苗,其具有来自流感H1N1和H3N2病毒的血凝素、基质蛋白1和核蛋白的mRNA,辅以cGAMP |
| 异源初免-加强方案中的ChAdOx1 NP+M1和MVA NP+M1 | 牛津大学詹纳研究所(英国) | 病毒载体 | 第一相 | 双剂量异源病毒载体构建体:修饰的痘苗病毒Ankara (MVA)和表达保守流感病毒抗原、核蛋白(NP)和基质蛋白-1的黑猩猩腺病毒ChAdOx2旨在刺激T细胞对流感病毒的反应。 Vatzia 2023 (PMID: 36792640), 福莱加蒂2022 (PMID: 35715352), Vatzia 2021 (PMID: 34804053), 考夫兰2018 (PMID: 29519670), 塔利2017 (PMID: 28724579), 安特罗布斯2014 (PMID: 24374965), 拉姆比青奥会(PMID: 23485942), NCT01818362, NCT01623518, 开发者网站 |
| ChAdOx1 NP+M1 prime,MVA-NP+M1 boost | 牛津大学詹纳研究所(英国)西奈山伊坎医学院(美国) | 病毒载体 | 临床前的 | 由ChAdOx1和MVA病毒载体疫苗递送的流感蛋白cHA、NP和M1旨在诱导有效的T和B细胞应答以提供广泛的保护。 |
| 茶毫克 | 中央研究院(台湾)、斯克里普斯研究所(美国)、国立阳明大学(台湾) | 重组蛋白质 | 临床前的 | 单糖基化嵌合HA (cHA毫克)具有共有H5头和共有H1茎抗原或C34-佐剂化的单糖基化HA茎(HA茎毫克);旨在通过CD4+和CD8+ T细胞应答引发针对一系列流感毒株和亚型的高滴度交叉反应性抗体。 |
| 嵌合细胞因子HA-VLP疫苗 | 日本Nerome生物资源研究所 | 病毒样颗粒(VLP) | 临床前的 | 包含嵌合细胞因子(CC)的VLPs,所述嵌合细胞因子包含M2蛋白、甲型流感神经氨酸酶柄 |
| 嵌合HA构建体 | 美国西奈山伊坎医学院 | 基于流感病毒 | 第一相 | 不同平台(包括LAIV和灭活裂解疫苗)与佐剂的顺序组合,或嵌合(cHA)或嵌合(mHA)构建体(cHA-LAIV-LAIV和cHA-LAIV-IIV,M2e)的顺序组合,由“外来的”HA头部结构域(来自禽流感病毒)和保守的茎结构域组成;具有不同头结构域和相同茎结构域的cHAs的顺序给药;旨在将体液免疫集中于高度保守的HA柄结构域。 (以前是: 基于cHA的LAIV组合) 埃德加2023 (PMID: 37871218), 普恩特-马萨格尔2023 (PMID: 37703367), 普恩特-马萨格尔2023 (PMID: 37342504), 米德2022 (PMID: 36533948), 艾迪罗2022 (PMID: 36496417), 朱2022(:35594401), 伊萨科娃-西瓦克2022 (PMID: 35461523), 福尔施韦勒2022 (PMID: 35461522), Guthmiller 2021, 纳克巴戈尔2021 (PMID: 33288923), 刘2021(:33440898), 伯恩斯坦2020 (PMID: 31630990), Destexhe 2020 (PMID: 32119974), 布勒克尔2019 (PMID: 31341648), 孙2019 (PMID: 31540436), 克拉梅尔2019 (PMID: 30715353), nachbaguer 2019(PMID:31839997), 刘2019(:31105689), Choi 2019 (PMID: 31032479), Sunwoo 2018 (PMID: 30223475), nachbaguer 2018(PMID:30044403), nachbaguer 2017(PMID:29263881), nachbaguer 2016(PMID:26719251), NCT03300050, NCT03275389, NCT02415842, 新闻稿2022年6月30日, 新闻稿2020年7月16日, 新闻稿2020年7月12日, 新闻稿2019年5月1日, 新闻故事2020年12月7日, NIH资助 |
| 嵌合HA蛋白 | 罗切斯特大学医学中心(美国) | 重组蛋白质 | 临床前的 | 新的嵌合HA蛋白(cH7/3)由来自季节性H3 HA的茎结构域和来自禽H7的头结构域组成。 |
| 嵌合M2e纳米组件 | 蒙特利尔魁北克大学(加拿大) | 非VLP纳米粒子 | 临床前的 | 使用嵌合M2e肽的基于纳米纤维的构建体。 |
| CIC疫苗 | 诺瓦瓦克斯(美国) | 非VLP纳米粒子 | 第二相 | 新型冠状病毒(新冠肺炎)和流感联合疫苗,使用新型冠状病毒病毒的全长稳定重组刺突蛋白(RS)和来自流感病毒的4种野生型重组血凝素(rHA)蛋白作为抗原。 马萨雷2021, NCT04961541, NCT05519839, 发展最新情况, 新闻稿2023年5月9日, 新闻稿2022年12月30日, 新闻稿2022年10月13日, 新闻稿2022年4月20日, 新闻稿2021年9月8日, 新闻稿2021年6月14日, 新闻稿2021年5月10日, 新闻故事2023年5月9日, Novavax演示2023, 开发者网站:Novavax |
| Clec9A-M2e | 新加坡国立大学(新加坡)、莫纳什大学(澳大利亚) | 重组蛋白质 | 临床前的 | 靶向策略,通过工程化重组抗Clec9A单克隆抗体,使M2e穿梭于特定的树突细胞亚群(cDC1 ),该抗体在其每条重链上与三个拷贝的M2e融合 |
| 中电大厦阻止 | 哥本哈根大学(丹麦)、斯克里普斯研究所(美国) | 病毒样颗粒(VLP) | 临床前的 | HAstem三聚体展示在刚性衣壳样颗粒(CLP)的表面,使用Tag/Catcher AP205平台。 |
| 眼镜蛇H1哈 | 北卡罗来纳大学(美国)、佐治亚大学(美国) | 重组蛋白质 | 临床前的 | 眼镜蛇HA疫苗与一种新型佐剂共同鼻内给药。佐剂包括PEI-Ox-DEX水凝胶、M7-CpG和cGAMP MPs 亨迪2023 (PMID: 37603310), 翁蒂韦罗斯-帕迪拉2023 (PMID: 37033992), 巴蒂2023 (PMID: 36436743), 巴蒂2022 (PMID: 35623484), 埃克什坦-李维2022 (PMID: 35704854), 瓦尔马2022 (PMID: 35353486) |
| 眼镜蛇-VLP | 佐治亚大学(美国)、克利夫兰诊所(美国) | 病毒样颗粒(VLP) | 临床前的 | 表达针对HA的COBRA(计算优化的广泛反应性抗原)的VLP构建体;旨在引发针对抗原性不同的流感病毒上的多个HA表位的高效价广泛交叉反应中和抗体。 卡洛克2023 (PMID: 37741893), 通用电气2023 (PMID: 36680243), 莫伊塞2022 (PMID: 35704783), 努内斯2021 (PMID: 34832509), 艾伦2021 (PMID: 34475872), 艾伦2021 (PMID: 33654128), 蕾妮儿2020 (PMID: 33115871), 黄2020(:31852790), 努内斯2020 (PMID: 31733946), 索托2020 (PMID: 31811019), 罗斯2019 (PMID: 30905528), 斯卡鲁普卡2019 (PMID: 31448974), Bar-Peled 2019 (PMID: 31481254), 艾伦2017 (PMID: 28789850), 黄2017 (PMID: 28978710), 卡特2017 (PMID: 28978709), 卡特2016 (PMID: 26912624), Crevar 2015 (PMID: 25671661), Giles 2012 (PMID: 22190399), Giles 2011 (PMID: 21320540), 新闻稿2023年9月20日, 新闻稿2022年7月14日, 开发者演示2023 |
| CodaVax | Codagenix(美国) | 基于流感病毒 | 第一相 | 通过合成减毒病毒工程(SAVE)产生的SAVE使用算法来“去优化”流感HA和NA基因片段,以减少在人类细胞中的翻译,导致病毒减毒,同时保留野生型病毒的免疫原性;旨在刺激针对来自多个季节和几十年的流感病毒(例如来自20世纪70年代和30年代的流感病毒)的免疫反应。 施陶夫特2019 (PMID: 31609986), 布罗德本特青奥会(PMID: 26655630), 杨2013(:23690603), 穆勒2010 (PMID: 20543832), NCT03926416, NCT05223179, NCT04146623, 新闻稿2022年6月14日, 开发者网站 |
| 组合聚合物纳米壳 | 台湾大学(台湾) | 非VLP纳米粒子 | 临床前的 | 具有旨在诱导T细胞应答的肽的共包封的纳米壳疫苗。 |
| 结合疫苗和P8载体肽 | TRIA生物科学(美国) | 非VLP纳米粒子 | 临床前的 | 基于纳米颗粒的肽,包含流感病毒ha干内高度保守的α螺旋结构域(用螺旋A表位修饰的肽);GLA-SE加入佐剂并旨在诱导针对小分子和基于蛋白质的表位的功能性抗体。 |
| 共轭透明质酸茎 | 墨尔本大学(澳大利亚) | 重组蛋白质 | 临床前的 | 衍生自PR8 HA头部结构域与HA茎蛋白的化学偶联肽。 |
| 保守T细胞表位DNA疫苗 | 圣路易斯大学(美国) | 基于核酸的 | 临床前的 | 包含多个pan-DR-和HLA-A2限制性高度保守流感抗原表位的t细胞靶向DNA疫苗。 状态:活动 |
| CTA1-3M2e-DD (FPM2e) | 哥德堡大学(瑞典)、根特大学(比利时) | 非VLP纳米粒子 | 临床前的 | 将基于M2e的融合蛋白与霍乱毒素衍生的佐剂(CTA1-DD)和脂质纳米颗粒组合;旨在增强抗M2e抗体和CD4+ T细胞反应。 奥莫坎耶2022 (PMID: 35260804), 贝纳斯科尼2020 (PMID: 32807838), 许塞克2020 (PMID: 31959882), 贝纳斯科尼2018年奥运会(PMID: 30271406), 埃利亚松2018 (PMID: 28295019), 埃利亚松2011 (PMID: 21481325), 埃利亚松2008 (PMID: 18243429) |
| ctLAIV | 延世大学(韩国) | 基于流感病毒 | 临床前的 | 胱天蛋白酶触发的减毒活流感疫苗(ctLAIV),其通过宿主胱天蛋白酶依赖性切割内部病毒蛋白而自我减毒。 |
| cVLPs | 嘉兴大学(中国) | 病毒样颗粒(VLP) | 临床前的 | 保守的柄结构域和4M2e共锚定到表面颗粒上,以产生嵌合流感病毒VLPs,旨在诱导体液和粘膜免疫应答。 |
| DCVC H1 HA mRNA-LNP | 美国国家过敏和传染病研究所 | 基于核酸的 | 第一相 | 编码甲型流感/加利福尼亚/07/2009 (H1N1)的全长HA的mRNA。 |
| 去糖基化改性透明质酸 | 厦门大学(中国) | 重组蛋白质 | 临床前的 | 来自H3N2的去糖基化修饰的HA蛋白,具有肌内施用的不同长度的聚糖 |
| 德尔塔流感 | 维瓦尔第生物科学公司(美国) | 基于流感病毒 | 第二相 | 自身佐剂性、非结构蛋白1 (NS1)缺陷型、复制缺陷型LAIV;以鼻腔喷雾剂的形式给药;旨在刺激干扰素、粘膜交叉中和IgA抗体、全身细胞毒性T细胞反应(Th1)和B细胞反应与交叉中和抗体和记忆T细胞反应。
尼科洛迪2019 (PMID: 31155415), 莫罗库蒂2014 (PMID: 24560674), 莫斯勒2013 (PMID: 24183981), 克伦2011 (PMID: 21490925), 瓦克2010 (PMID: 20039806), NCT01078701, NCT03745274, NCT00724997, 新闻稿2023年6月5日, 新闻稿2023年3月29日, 新闻稿2021年11月29日, 新闻稿2020年11月23日, 新闻稿2019年6月19日, 新闻稿2018年4月4日, 新闻稿2018年2月12日, 开发者网站, NIH资助 |
| 具有编码保守抗原的质粒的DNA构建体 | 国家病毒学和生物技术研究中心(俄罗斯) | 基于核酸的 | 临床前的 | 编码基于HA柄和M2蛋白的人工抗原的DNA,以诱导交叉保护性体液和细胞介导的反应。 |
| DNA主亚单位蛋白增强剂 | 上海生物制品研究所(中国) | 基于核酸的 | 临床前的 | 编码保守M1 (prime)和可溶性重组M1亚基蛋白(boost)的DNA质粒;旨在诱导细胞和体液免疫反应,以针对异亚型病毒感染进行交叉保护。 |
| 双层蛋白质纳米颗粒 | 佐治亚州立大学(美国) | 非VLP纳米粒子 | 临床前的 | 表达多价保守抗原(M2e、NP、HA茎结构域)的层状蛋白质纳米颗粒;旨在诱导广泛的交叉保护性免疫反应。 状态:活动 王2023(:36265560), 宋2022(:35810540), 董2022(:35084819), 董2022, 马2021(:34743020), 王2021(:34179728), 唐2020(:33114336), 王2020(:31840437), 邓2018(:30065113), 邓2018(:30394725), 王2018(:30365905), 邓2018(:29367723), 昌2018(:30092060), 邓2017(:28622575), 王2014(:23988715), 新闻稿2019年7月1日, NIH资助 |
| UFV-001 | 紧急生物解决方案(美国) | 非VLP纳米粒子 | 第一相 | 基于纳米颗粒的疫苗,在生产过程中自组装,并显示出针对甲型流感病毒1组和2组的交叉反应性HA抗原。自组装HA稳定干细胞纳米粒子技术由NIAID开发并获得授权。UFluA由DP-UFluA (1:1的A1-ssnp和A2-ssnp抗原)组成,并含有氢氧化铝和CpG佐剂。 |
| EFn-3xM2e-HA2+PA | 德克萨斯理工大学(美国) | 重组蛋白质 | 临床前的 | 由3个串联M2e重复加上HA2组成的抗原,与去毒炭疽水肿毒素(EFn加上PA)组合;旨在引发抗原特异性IgG反应;鼻内给药。 |
| 流感病毒mRNA | CureVac(德国)、GSK(美国) | 基于核酸的 | 第二相 | 修饰的(Flu-SV-mRNA)和未修饰的(CVSQIV)基于mRNA的脂质纳米粒。Flu-SV-mRNA是表达H1N1抗原的单价构建体。 鲁兹2017 (PMID: 29263884), 卡伦2013 (PMID: 23921513), Petsch 2012 (PMID: 23159882), NCT05252338, NCT05446740, NCT05823974, 新闻稿2023年11月14日, 新闻稿2023年11月1日, 新闻稿2023年9月12日, 新闻稿2023年8月5日, 新闻稿2023年4月25日, 新闻稿2023年1月30日, 新闻稿2022年10月2日, 葛兰素史克演示2023, 开发者网站:CureVac, 开发者网站:GSK |
| 流感病毒 | ConserV生物科学(英国),Imutex(英国) | 重组蛋白质 | 第二相 | 源自内部蛋白质(M1、IAV-NP、IBV-NP和M2)保守区的基于肽的构建体,旨在通过细胞毒性T细胞应答清除病毒,提供针对甲型和乙型流感的广泛保护性免疫应答。 Oftung 2022 (PMID: 36146606), Pleguezuelos 2020 (PMID: 32150750), Pleguezuelos 2020 (PMID: 32194999), 阿巴西2020 (PMID: 32286631), 范多恩2017 (PMID: 28376743), Pleguezuelos 2015 (PMID: 26084515), Pleguezuelos 2015 (PMID: 25994549), Pleguezuelos 2012 (PMID: 22575166), 斯托洛夫2007 (PMID: 17668898), 新闻稿2020年3月16日, 新闻稿2019年1月11日, NCT 02962908/EudraCT 2015-001932-38, NCT03180801, NCT01226758, NCT01181336, 开发者网站:ConserV Bioscience, 开发者网站:Imutex, NIH资助 |
| FluMos自组装纳米粒子 | 美国国家过敏和传染病研究所 | 非VLP纳米粒子 | 第一相 | 使用计算设计的纳米粒子免疫原,其在自组装蛋白质纳米粒子上以有序阵列可控地展示不同的HA三聚体;旨在引发针对异亚型流感病毒的HAI活性和保护性干细胞定向中和抗体。 (原:FluMos-v1) 阿拉班扎2023 (PMID: 37451877), 杨2023(:36906406), 沙贾汗2023 (PMID: 36723411), 博约格鲁-巴纳姆2021 (PMID: 33762730), Kanekiyo 2019 (PMID: 30742080), 克拉默2019评论(PMID: 30742079), 乔尔杰夫2018年奥运会(PMID: 29451984), Kanekiyo 2013 (PMID: 23698367), NCT05968989, NCT04896086, 新闻故事2023年9月27日, 新闻稿2023年9月15日, 新闻稿2021年6月1日, 新闻稿2021年5月6日, NIH资助, NIH资助, 开发者网站:NIAID |
| FP-01.1 | 免疫靶向系统(Altimmune)(英国) | 重组蛋白质 | 第二相 | 由连接到惰性氟碳链的6个合成肽组成的新构建体,包封多个保守表位(NP、M和PB1/PB2);旨在产生T细胞对不同流感毒株的反应。 弗朗西斯2015 (PMID: 24928790), NCT01265914, NCT02071329, NCT01677676, NCT01701752, 开发者网站 |
| G1 mHA | 让桑疫苗和预防,J&J(荷兰) | 重组蛋白质 | 第二相 | 基于干细胞的第1组HA抗原,基于去除免疫显性HA头部区域同时稳定干细胞区域结构的方法;旨在诱导广泛中和的抗体。 (原:迷你哈) 斯瓦特2023 (PMID: 37952003), Kil 2019 (PMID: 31725776), 范德鲁比2018 (PMID: 29977611), 范德鲁比2018 (PMID: 30369928), Impagliazzo 2015 (PMID: 26303961), NCT05901636, 开发者网站 |
| 伽马氟 | 伽马疫苗(澳大利亚) | 基于流感病毒 | 临床前的 | 用γ射线灭活的全流感病毒,以防止病毒复制,保持外部和内部蛋白抗原完整;旨在刺激抗体和细胞毒性T细胞以提供交叉保护性免疫;鼻内给药。 单身2021 (PMID: 34899711), 大卫2017 (PMID: 28109709), 扶余拉2010 (PMID: 20164231) |
| 糖脂肽 | 莫纳什大学(澳大利亚)、惠灵顿维多利亚大学(新西兰) | 重组蛋白质 | 临床前的 | 来自免疫原性MHC类结合CD8+ T细胞表位的合成长肽(SLP ),其共价连接到α-半乳糖神经酰胺佐剂(一种自然杀伤T细胞激动剂);旨在刺激流感特异性记忆细胞毒性T淋巴细胞和针对保守IAV表位的交叉保护性免疫反应。 |
| H1马赛克HA | 内布拉斯加大学林肯分校(美国),圣裘德儿童研究医院(美国) | 病毒载体 | 临床前的 | 基于全长H1 HA或多价HA共有基因的复制缺陷型腺病毒5型载体镶嵌HA;旨在诱导针对不同流感毒株的细胞和体液免疫反应。 科德2019 (PMID: 31771231), 林格尔2017 (PMID: 29097763), 威比2015 (PMID: 26469190) |
| H1,H3眼镜蛇IIV | 佐治亚大学(美国)、克利夫兰诊所(美国) | 基于流感病毒 | 临床前的 | 表达H1和H3计算优化的广泛反应性抗原(眼镜蛇)HA抗原的重配流感病毒。 Dzimianski 2023 (PMID: 37185989), 长岛2023 (PMID: 36851561), 罗斯2022 (PMID: 36016202), 阿巴迪2022 (PMID: 35916534), 长岛2022 (PMID: 35697384), 艾伦2022 (PMID: 35289635), 黄2021(:34358209), 索托2021 (PMID: 33853960), 索托2018 (PMID: 31022693), 艾伦2018 (PMID: 30265682) |
| 总部位于H1的cHAs | 加纳大学(加纳) | 重组蛋白质 | 临床前的 | 嵌合HAs (cHAs)通过构建一致序列产生,旨在诱导交叉反应性抗体。 |
| h1c mRNA-LNP | 联合疫苗国家工程技术研究中心(中国) | 基于核酸的 | 临床前的 | 一种脂质纳米粒(LNP)包裹的核苷修饰的mRNA疫苗(mRNA-LNPs ),编码一个共有的全长HA序列(H1c) |
| H2孔 | 内布拉斯加大学林肯分校(美国) | 病毒载体 | 临床前的 | 针对H2甲型流感病毒的腺病毒载体集中式共有HA构建体,其设计为当在初始-加强策略中首先由Ad6接着由Ad5递送时,诱导广泛的保护性抗体滴度。 |
| HA迷你茎 | 香港大学(中国香港特别行政区) | 重组蛋白质 | 临床前的 | 基于H5的HA小茎蛋白折叠为三聚体在融合前确认HA茎;使用四分之一全长HA将免疫反应集中于HA茎结构域中已知广泛中和抗体的保守表位。 |
| 透明质酸干蛋白 | 美国国家过敏和传染病研究所(NIAID)、印度科学研究所(印度) | 重组蛋白质 | 临床前的 | 构建HA干片段免疫原的新方法:旨在克服HA头部免疫优势和刺激抗干细胞生发中心B细胞和抗体反应。 安杰莱蒂2019 (PMID: 31213541), 萨顿2017 (PMID: 29263889), 马拉乔苏拉2015 (PMID: 26167164), 马拉约苏拉2014 (PMID: 24927560), 博马坎蒂2012 (PMID: 23015722) |
| HA三聚体 | 加州理工学院(美国) | 非VLP纳米粒子 | 临床前的 | 使用SpyCatcher-SpyTag“插入和展示”策略将三聚体HA共价偶联到具有不同数量附着位点的对称颗粒上,从而产生展示来自多达8种1型和2型甲型流感病毒毒株的HA抗原的镶嵌颗粒。 |
| HA-钳夹蛋白质类 | 昆士兰大学(澳大利亚) | 重组蛋白质 | 临床前的 | 分子钳,其利用高度稳定的三聚化结构域来允许构象稳定的灌注血凝素HA的有效生产和纯化 |
| HA-F DNA疫苗 | 吉林大学(中国) | 基于核酸的 | 临床前的 | 通过将流感病毒HA与自组装铁蛋白纳米颗粒融合而设计的DNA疫苗旨在提供强大的免疫原性、高保护效力,并且是可快速生产的有效疫苗 |
| 透明质酸脂质体 | 莫纳什大学(澳大利亚) | 非VLP纳米粒子 | 临床前的 | 具有在脂质体表面功能化的HA免疫原的脂质纳米粒亚单位疫苗的生产;使用先进的微流体混合技术(NanoAssemblr ),能够快速制备脂质体表面HA抗原浓度增加的脂质体疫苗;旨在引发透明质酸特异性B细胞和滤泡辅助性T细胞免疫反应。 |
| HA-T-AGM蛋白 | 转化健康科技研究所(印度) | 重组蛋白质 | 临床前的 | 一种在哺乳动物Expi293F表达系统中表达的重组全长可溶性三聚体HA蛋白,经皮内给药并使用Addavax佐剂 |
| 哈-VLP-中青旅 | 佐治亚州立大学(美国) | 病毒样颗粒(VLP) | 临床前的 | 旨在诱导细胞和体液免疫的细胞因子佐剂的流感血凝素病毒样颗粒(HA-VLP)疫苗 |
| HA/GP纳米粒子 | 佐治亚州立大学(美国) | 非VLP纳米粒子 | 临床前的 | 聚乙烯亚胺功能化的氧化石墨烯纳米粒子(GP纳米粒子);鼻内给药;旨在增强HA免疫原性 |
| 哈/M1·VLP | 佐治亚州立大学(美国) | 病毒样颗粒(VLP) | 临床前的 | 连续施用含有来自HA系统发育组1的H1、H8和H13,或来自HA系统发育组2的H3、H4和H10,或各种组合的异亚型HA VLPs旨在引发针对同一HA系统发育群或两个群中的不同病毒的广泛保护。 |
| ha2ha-铁蛋白纳米颗粒 | 美国国家过敏和传染病研究所 | 非VLP纳米粒子 | 第一相 | 铁蛋白纳米颗粒疫苗由来自H2N2疫情毒株A/Singapore/1/57的HA的胞外域与来源于幽门螺杆菌其在纳米颗粒表面上以有序的阵列显示了八个抗原性完整的H2流感病毒HA三聚体 杨2023(:37781590), 尼尔森2022 (PMID: 36289232), 豪泽2022 (PMID: 35115706), 安德鲁斯2022 (PMID: 35115707), 凯利2020 (PMID: 32434990), 2018年的达里卡尔(PMID: 30185594), NCT03186781, 新闻稿2022年5月11日, 开发者网站:NIAID |
| HAdV-5-题词 | 内布拉斯加大学林肯分校(美国) | 病毒载体 | 临床前的 | 计算设计为通用人H3流感疫苗,由三种H3病毒组成,在复制缺陷型腺病毒5型载体(HAdV-5)中表达。 |
| HAsd蛋白 | 西南交通大学医学院(中国) | 重组蛋白质 | 临床前的 | 以乳酸乳球菌表面展示的HA柄结构域(HAsd)为目标的重组蛋白为临床前的构建体;旨在提供针对不同甲型流感病毒的交叉免疫。 |
| HBc VLPs | 中国科学院(中国) | 病毒样颗粒(VLP) | 临床前的 | 使用乙型肝炎病毒核心(HBc) VLP,用内部NP和外部M2e抗原仿生双抗原VLP构建体;旨在诱导交叉保护性免疫反应。 |
| HBc-4M2eh (Uniflu) | 俄罗斯科学院(俄罗斯)、VA Pharma(俄罗斯) | 病毒样颗粒(VLP) | 第一相 | 包含融合在乙型肝炎病毒核心抗原(HBc)的免疫显性环内的4个拷贝的M2e的疫苗;旨在诱导M2e抗体反应。 (原:M2e VLP) 梅真斯卡娅2019 (PMID: 31629405), 齐巴洛娃2015 (PMID: 25976545), 瑞文尼青奥会(PMID: 25937448), 瑞文尼青奥会(PMID: 22339631), NCT03789539 |
| 螺旋-茎纳米颗粒 | 美国国家过敏和传染病研究所 | 非VLP纳米粒子 | 临床前的 | 基于纳米颗粒的疫苗,每个纳米颗粒亚单位显示保守流感螺旋A区的多个拷贝 |
| HMN-PP | 吉林大学(中国) | 非VLP纳米粒子 | 临床前的 | 多表位纳米疫苗,MHF,由HA2、M2e和NP的保守线性表位组成,融合到P结构域的三个表面环或铁蛋白 |
| 杂交融合蛋白组合疫苗 | 埃默里大学(美国)、佐治亚州立大学(美国) | 病毒样颗粒(VLP) | 临床前的 | 流感和新型冠状病毒(新冠肺炎)VLP联合疫苗,使用流感病毒样颗粒(VLP)通过蛋白转移与融合于GM-CSF的糖基磷脂酰肌醇(GPI)锚定的新型冠状病毒RBD融合作为佐剂 |
| IAV-纳诺瓦 | 爱荷华大学(美国),爱荷华州立大学(美国) | 非VLP纳米粒子 | 临床前的 | 基于五嵌段共聚物胶束和聚酸酐纳米颗粒的组合纳米疫苗平台,整合了来自H1N1的血凝素(HA)和核蛋白(NP)抗原;旨在增强细胞介导的免疫反应。 状态:活动 罗斯2019 (PMID: 30663733), 撒迦利亚2018 (PMID: 30233573), 授权摘要, NIH资助, 专利 |
| 鼻内NLP:NP | 罗切斯特大学医学中心(美国) | 非VLP纳米粒子 | 临床前的 | 用佐剂连接到NP的自组装纳米脂蛋白颗粒 |
| 倒置的哈VLP | 美国佐治亚理工学院,美国西奈山伊坎医学院 | 病毒样颗粒(VLP) | 临床前的 | 一种展示多拷贝抗原结合片段(Fab)的VLP,该片段识别HA头部的顶点,引起随后的HA结合,导致其以反向呈递。 |
| 含ISCOMs/MPLA佐剂的SDAD蛋白纳米颗粒 | 佐治亚州立大学(美国) | 非VLP纳米粒子 | 临床前的 | 由流感核蛋白(NP)作为核心和NA1-M2e或NA2-M2e融合蛋白作为包被抗原组成的核/壳蛋白纳米颗粒,通过免疫刺激复合物(ISCOMs单磷酰脂质A (MPLA)佐剂促进的SDAD异双功能交联 |
| LAIV+4M2e | 实验医学研究所(俄罗斯) | 基于流感病毒 | 临床前的 | 表达额外4个M2e串联重复序列(4 M2e)的重组LAIV;旨在增强M2e介导的针对异源病毒的交叉保护。 梅真斯卡娅2021 (PMID: 34209093), 梅真斯卡娅2021 (PMID: 33535408), Kotomina 2020 (PMID: 33153089) |
| LHNVD-105 | 长角牛疫苗和诊断(美国) | 重组蛋白质 | 临床前的 | 靶向HA、NA和M2e的多表位非偶联和CRM偶联肽;用ALFQ(陆军脂质体)佐剂配制 Rikhi 2023 (PMID: 37766144), Sei 2021 (PMID: 34202178), Sei 2018, 新闻稿2023年9月27日, 新闻稿2022年10月24日, 开发者网站:长角牛疫苗和诊断 |
| 脂质体肽疫苗 | 格里菲斯大学(澳大利亚) | 非VLP纳米粒子 | 临床前的 | 脂质体疫苗辅以TLR4激动剂3D-PHAD,表达保守的甲型流感M2e表位 |
| m-cNA-M2e VLP | 佐治亚州立大学(美国) | 病毒样颗粒(VLP) | 临床前的 | 一种表现出肌内施用的共有多神经氨酸酶(NA)亚型(cN1、cN2、BcNA)和M2胞外域(M2e)串联重复序列(mcNA-M2e VLP)的VLP实体。 费兰米2022 (PMID: 36252577), 金2022 (PMID: 36006890), 新闻稿2022年8月25日 |
| 基于M2e的重组融合蛋白 | 俄罗斯卫生部(俄罗斯) | 重组蛋白质 | 临床前的 | 基于M2e的4个串联拷贝和HA2的保守片段的组合的重组植物产生的蛋白质(Flg4M2eHA2-1 ),融合到细菌鞭毛蛋白作为粘膜免疫的佐剂;鼻内给药。 斯捷潘诺娃2022 (PMID: 36560509), 布洛赫纳2020 (PMID: 32013187), 齐巴洛娃2018 (PMID: 30138320), 斯捷潘诺娃2018年奥运会(PMID: 29631629), 斯捷潘诺娃2018年奥运会(PMID: 29713522), 马尔达诺瓦2016 (PMID: 26710263), 马尔达诺瓦2015 (PMID: 26022390), 斯捷潘诺娃青奥会(PMID: 25799221) |
| M2e纳米团簇 | 德克萨斯理工大学(美国)、乔治亚州立大学(美国) | 非VLP纳米粒子 | 临床前的 | 无添加剂交联M2e纳米团簇(NCs) 状态:活动 内索维奇2023 (PMID: 37455612), 威尔克斯2022 (PMID: 36346898), 威尔克斯2021 (PMID: 34432416), 英欧尔2021 (PMID: 34301431), 比姆勒2019 (PMID: 31507643), 陶2017(:28161578), 陶2015(:25842219), 陶2014(电话:23829488), NIH资助 |
| M2e VLP议员 | 美世大学(美国) | 病毒样颗粒(VLP) | 临床前的 | 包封在微粒基质中的含佐剂的M2e基VLP Braz Gomes 2023 (PMID: 37445784), Braz Gomes 2022 (PMID: 36146733), Braz Gomes 2022 (PMID: 35304243), Braz Gomes 2021 (PMID: 34835255) |
| M2e基自组装纳米粒子 | 复旦大学(中国) | 非VLP纳米粒子 | 临床前的 | 携带M2e表位的自组装纳米颗粒,旨在诱导针对不同流感亚型的M2e抗体。 |
| 基于M2e的VLPs (ACAM-FluA) | 根特大学(比利时)、赛诺菲巴斯德(美国) | 病毒样颗粒(VLP) | 第一相 | 基于M2e的VLP构造;旨在诱导交叉反应性CD8+ T细胞应答。 Schotsaert 2016 (PMID: 27072615), 依班娜青奥会(PMID: 23527091), 德菲莱特2008 (PMID: 18835315), NCT00819013, 开发商网站:赛诺菲 |
| M2e-CRM197轭合物 | 中国科学院(中国) | 重组蛋白质 | 临床前的 | M2e肽与CRM的双位点特异性结合197变性中的载体蛋白。 |
| M2e-H3梗 | 佐治亚州立大学(美国) | 重组蛋白质 | 临床前的 | 嵌合M2e和H3血凝素(HA)茎蛋白疫苗,通过将M2e重复序列与具有稳定HA1 N-和C-末端区域和点突变的工程化H3茎结构域遗传连接而工程化 |
| M2e/CpG-ODN/TMC | 农业研究、教育和推广组织(伊朗) | 非VLP纳米粒子 | 临床前的 | 用封装在N-三甲基壳聚糖(TMC)纳米颗粒中的M2e/CpG-ODN抗原进行鼻接种 |
| M2e5x VLP(多种方法) | 佐治亚州立大学(美国),庆熙大学(韩国) | 病毒样颗粒(VLP) | 临床前的 | 单独或与其他技术组合的保守串联重复M2e5x VLPs,包括:HA VLPs、鞭毛蛋白VLPs、微针贴片IIV和鼻内给药;旨在诱导异亚型交叉保护性免疫。 康2019(:31246961), 李2019 (PMID: 30685658), 金2019 (PMID: 31003421), 金2018 (PMID: 30199754), 金2018 (PMID: 30241300), 李2018 (PMID: 29324805), 金2017 (PMID: 28887040), 金2014 (PMID: 25171841), 金2013 (PMID: 23247101) |
| M2SR | 弗卢根(美国) | 基于流感病毒 | 第二相 | 基于不表达M2离子通道蛋白(M2缺陷型)的野生型流感病毒的用于甲型或乙型流感病毒的新型单一复制(SR)平台;鼻内给药;旨在引发针对保守HA干细胞和全身及粘膜免疫反应的交叉反应性抗体,从而阻断病毒在肺中的复制并提供针对流感病毒的跨谱系保护。这种候选疫苗的四价形式Quad M2SR目前正处于临床前开发阶段。
希尔-巴托尔斯基2023 (PMID: 37112710), 艾登2023 (PMID: 36350017), 萨瓦2022 (PMID: 36560540), 艾登2022 (PMID: 34323977), 艾登2021, Moser 2019 (PMID: 31280945), 哈达2018 (PMID: 30007825), 哈达2017 (PMID: 28668565), 萨拉瓦尔2016 (PMID: 27595896), 哈达2011 (PMID: 21272601), 渡边2009 (PMID: 19321619), 新闻故事2022年3月10日, 新闻稿2022年9月11日, 新闻稿2022年9月7日, 新闻稿2022年8月1日, 新闻稿2022年7月27日, 新闻稿2022年6月22日, 新闻稿2021年8月2日, 新闻稿2021年7月1日, 新闻稿2021年5月25日, 新闻稿2019年8月28日, EudraCT 2017-004971-30, NCT03999554, NCT03553940, NCT02822105, NCT04785794, NCT04960397, NCT05163847, 开发者网站 |
| MAV-1公顷 | 列日大学(比利时) | 病毒载体 | 临床前的 | 口服施用的表达the HA的重组小鼠腺病毒1型(MAV-1) |
| 微共有DNA疫苗 | 威斯特研究所(美国) | 基于核酸的 | 第一相 | 基于4种编码H1和H3 HA抗原的质粒的合成微共识DNA平台,每种抗原包含基于H1和H3 HA抗原一级序列子集分析的有限共识序列;旨在诱导针对H1N1和H3N2流感不同分离株的HAI滴度的保护水平;通过皮内/肌内电穿孔(EP)递送;旨在引发抗体和T细胞反应。 徐2020(:32328416), 颜2018(:29100705), 艾略特2018 (PMID: 30062926), 颜2014(:24631084), Kichaev 2013 (PMID: 23954979), NCT01403155, NCT01405885, NCT01142362, NCT01184976, 开发者网站 |
| 迷你HA-LS纳米疫苗 | 华中农业大学(中国) | 非VLP纳米粒子 | 临床前的 | 通过SpyTag/SpyCatcher在lumazine合酶(LS)纳米颗粒上表达的Mini-HA蛋白旨在引发有效的免疫反应 |
| MLN mRNA | 上海生物制品研究所(中国) | 基于核酸的 | 临床前的 | 一种新的基于mRNA的多抗原流感疫苗,其基于单个mRNA分子与甲型流感病毒三种保守抗原的串联,包括M2离子通道(M2e)的胞外域、血凝素柄区(LAH)的长α螺旋和核蛋白(NP) |
| 改良mRNA疫苗 | 辉瑞(美国)、生物科技(德国) | 基于核酸的 | 第三阶段 | 编码世卫组织推荐毒株的下一代mRNA季节性流感疫苗 多尔金2021 (PMID: 34635829), 阿巴西2021 (PMID: 34751730), 新闻稿2023年10月31日, 新闻稿2022年9月14日, 新闻稿2022年9月14日, 新闻稿2021年9月27日, 新闻稿2018年8月16日, NCT05540522, NCT05052697, 辉瑞2022年第二季度收益电话会议, 开发者网站:辉瑞, 开发者网站:BioNTech |
| 基于modRNA的组合 | 辉瑞(美国)、生物科技(德国) | 基于核酸的 | 第二相 | 组合mRNA新型冠状病毒(新冠肺炎)和流感疫苗,其包含编码新型冠状病毒野生型刺突蛋白和Omicron BA.4/BA.5亚变异体的刺突蛋白的mRNA株以及编码四种不同流感毒株的血凝素的mRNA链 NCT05596734, 新闻稿2023年10月26日, 新闻稿2022年9月12日, 新闻稿2022年3月11日, 开发者网站:辉瑞, 开发者网站:BioNTech |
| 镶嵌和嵌合HA | 美国西奈山伊坎医学院 | 重组蛋白质 | 临床前的 | 重组镶嵌和嵌合HA的新构建体,表达为可溶性三聚体蛋白;旨在诱导针对甲型和乙型流感病毒株的广泛保护性免疫反应。 鲁比杜2021 (PMID: 33593972), 孙2019 (PMID: 30944178), 克拉梅尔2019 (PMID: 30715353), 厄姆勒2017 (PMID: 28356526), 蛋墨2014 (PMID: 24155380), 玛吉2013 (PMID: 23903831) |
| 基于镶嵌HA的全灭活病毒 | 美国西奈山伊坎医学院 | 基于流感病毒 | 临床前的 | 活化的乙型流感病毒展示镶嵌HA (mHA)蛋白,将免疫反应导向HA的免疫亚优势保守表位。 |
| 马赛克VLPs | 中山大学(中国) | 病毒样颗粒(VLP) | 临床前的 | 一种HA和NA镶嵌免疫原混合物,包含大多数表现为病毒样颗粒(VLPs)的人类季节性流感病毒(H1N1和H3N2)的潜在T细胞表位 |
| mRNA构建体 | 赛诺菲巴斯德(美国) | 基于核酸的 | 第二相 | 单价和四价mRNA疫苗构建体;两个候选人的区别仅在于含有mRNA的LNP 以前:mRNA 奇武库拉2021 (PMID: 34916519), 多尔金2021 (PMID: 34635829), NCT05553301, NCT05426174, NCT05624606, NCT05650554, NCT05829356, NCT06118151, 赛诺菲投资者演示, 新闻稿2021年6月22日, 开发商网站:赛诺菲 |
| mRNA-1010 | 现代(美国) | 基于核酸的 | 第三阶段 | 基于修饰mRNA的脂质纳米颗粒候选疫苗mRNA-1010(在第3阶段),一种四价制剂,含有来自四种季节性流感毒株(A/H1N1、A/H3N2和B/Yamagata-和B/Victoria-谱系)的HA抗原 李2023 (PMID: 37336877), 巴林2022, 多尔金2021 (PMID: 34635829), 费尔德曼2019 (PMID: 31079849), 巴林2017 (PMID: 28457665), 梁2017(:28958578), 林德格伦2017 (PMID: 29181005), NCT05566639, NCT05827978, NCT05415462, NCT05868382, NCT05606965, NCT05397223, NCT04956575, NCT03076385, NCT03345043, 开发者网站, 新闻稿2023年9月13日, 新闻稿2023年3月8日, 新闻稿2023年4月5日, 新闻稿2023年4月11日, 新闻稿2023年2月16日, 新闻稿2022年9月8日, 新闻稿2022年6月7日, 新闻稿2022年4月11日, 新闻稿2022年3月24日, 新闻稿2021年12月10日, 新闻稿2021年7月7日, 现代展示2022 |
| mRNA 1011和mRNA 1012 | 现代(美国) | 基于核酸的 | 第二相 | 修饰的mRNA脂质纳米粒mRNA 1011/1012其为季节性五价/六价疫苗候选物,包含更多HA抗原(例如H3、H1)以扩大毒株匹配 巴林2017 (PMID: 28457665), 梁2017(:28958578), 林德格伦2017 (PMID: 29181005), 新闻稿2023年4月11日, NCT05827068, 开发者网站 |
| mRNA 1020和mRNA 1030 | 现代(美国) | 基于核酸的 | 第二相 | 修饰的mRNA脂质纳米粒mRNA 1020和mRNA 1030其掺入HA和NA抗原以靶向病毒的更保守区域 巴林2022, 多尔金2021 (PMID: 34635829), 巴林2017 (PMID: 28457665), 梁2017(:28958578), 林德格伦2017 (PMID: 29181005), NCT05333289, 开发者网站, 新闻稿2023年4月11日, 新闻稿2023年2月16日 |
| mRNA-1083 | 现代(美国) | 基于核酸的 | 第三阶段 | 新型冠状病毒(新冠肺炎)和流感联合疫苗。编码4种流感病毒株的新冠肺炎刺突蛋白和流感HA糖蛋白的单剂量mRNA疫苗。 (以前:mRNA-1073和mRNA-1083) NCT06097273, NCT05827926, NCT05375838, 新闻稿2023年10月24日, 新闻稿2023年4月10日, 现代展示2021年4月11日, 新闻故事2021年9月10日, 新闻稿2021年9月9日, 新闻故事2023年10月26日, 开发者网站:Moderna |
| mRNA-1230 | 现代(美国) | 基于核酸的 | 第一相 | 肌肉注射多组分mRNA新型冠状病毒(新冠肺炎)、流感和呼吸道合胞病毒(RSV)疫苗。 |
| mRNA流感 | 国家公共卫生和环境研究所(荷兰) | 基于核酸的 | 临床前的 | 编码H1N1流感病毒三个保守内部蛋白的核苷修饰的mRNA-LNP,NP,M1和PB1 |
| mRNA-LNP | 美国宾夕法尼亚大学西奈山伊坎医学院 | 基于核酸的 | 临床前的 | 核苷修饰的mRNA疫苗构建体编码来自所有20种已知甲型和乙型流感病毒亚型的保守抗原或血凝素抗原,旨在诱导抗原特异性细胞和体液免疫应答,以保护抵抗不同的流感病毒株。 状态:活动 柯比2023 (PMID: 36708722), 新镇2023 (PMID: 36631700), Gouma 2022 (PMID: 36533954), 佩埃塔2022, 奥利里2022 (PMID: 36450841), 开尔文2022 (PMID: 36423290), 阿雷巴洛2022 (PMID: 36423275), 麦克马洪2022 (PMID: 36322769), 帕迪2022 (PMID: 35945226), 村松2022 (PMID: 35131437), 阿拉梅2021, 弗雷恩2021 (PMID: 34485597), 威利斯2020 (PMID: 31915303), 弗雷恩2020 (PMID: 32359470), Pardi 2018 (PMID: 30135514), Limberis 2013 (PMID: 23720583), 新闻稿2022年6月12日, 新闻稿2022年6月12日, 新闻稿2022年11月25日, NIH资助, NIH资助 |
| mRNA/LNP疫苗 | 默克公司(美国) | 基于核酸的 | 临床前的 | 编码各种形式流感抗原的LNP包被的化学修饰的mRNA疫苗 |
| 多抗原DNA疫苗 | 华盛顿大学(美国) | 基于核酸的 | 临床前的 | 含佐剂的多抗原DNA疫苗,由HA、M2和NP抗原组成,分3次连续给药;旨在诱导粘膜反应和交叉反应性T细胞反应。 |
| 基于多表位广告 | 普渡大学(美国) | 病毒载体 | 临床前的 | 使用H5N1流感病毒的相对保守的免疫原性结构域的基于腺病毒载体的多表位构建体:M2胞外域(M2e)、血凝素(HA)融合域(HFD)、核蛋白的T细胞表位(TNP)和HA α螺旋域(HαD);旨在诱导体液和细胞介导的免疫反应。 |
| 多聚体-001 (M-001) | BiondVax制药公司(以色列) | 重组蛋白质 | 第三阶段 | 9个保守的M1、NP和HA表位的单一重组蛋白;旨在诱导T细胞应答和增强B细胞对循环和新菌株的应答。 阿特马尔2023 (PMID: 36941155), 菲利普森2019 (PMID: 35174313), 洛厄尔2017 (PMID: 28065476), 范多恩2017 (PMID: 28296763), 阿斯特蒙2014 (PMID: 25173483), 戈特利布2014 (PMID: 25172355), 阿斯特蒙2012 (PMID: 22318394), NCT03450915, NCT03058692, NCT02691130, NCT02293317, NCT01419925, NCT01146119, NCT01010737, NCT00877448, 新闻稿2020年10月23日, 开发者网站:现在称为Scinai免疫疗法 |
| 多重HA-DNA | 奥斯陆大学(挪威) | 基于核酸的 | 临床前的 | 靶向APCs的DNA编码疫苗蛋白;使用NA的胞外域作为抗原或来自6种1组流感病毒(H5、H6、H8、H9、H11和H13)中每一种的HA基因,并将它们插入到DNA疫苗形式中;旨在诱导NA免疫并将HA蛋白抗原递送至APCs上的MHC II类分子。 施耐德2023 (PMID: 37201525), 韦宁豪斯2023 (PMID: 36926694), 安徒生2021 (PMID: 34691056), 格罗德兰2020 (PMID: 32269566), 2020年雅典奥运会(PMID: 32128342), 安德森2018 (PMID: 29427414) |
| 多价DNA纳米疫苗 | 台州大学(中国) | 非VLP纳米粒子 | 临床前的 | pβH7N2SH9/DGL纳米粒子是通过将pβH7N2SH9封装在树状接枝聚l-赖氨酸纳米粒子中而开发的 |
| MVA-NP | 德国感染研究中心(DZIF)(德国) | 病毒载体 | 临床前的 | 重组MVA候选疫苗,其在五种新设计的嵌合痘病毒启动子的转录控制下递送高度保守的IAV内部核蛋白(NP ),以进一步增加重组病毒(MVA-NP)的免疫原性 |
| MVA-NP+M1 (VTP-100) | 真空技术公司(英国) | 病毒载体 | 第二相 | 改良的安卡拉痘苗病毒(MVA)-表达核蛋白(NP)和M1蛋白(MVA-NP+M1)的载体构建体;靶向T细胞对流感病毒核蛋白和基质1核心蛋白的反应;与许可的四价灭活流感病毒疫苗共同施用。 埃文斯2022 (PMID: 35305317), 瓦尔肯堡2022 (PMID: 35305315), 巴特勒2021 (PMID: 34451976), Puksuriwong 2020 (PMID: 31740938), 斯威兹2019 (PMID: 32089822), 福勒加蒂2019 (PMID: 30909516), 马林2016 (PMID: 26902548), 安特罗布斯2014 (PMID: 23831594), 穆拉基青奥会(PMID: 23589155), 鲍威尔2013 (PMID: 23658773), 莉莉艾青奥会(PMID: 22441650), 安特罗布斯2012 (PMID: 23118984), 贝尔图德2011 (PMID: 21148512), 新闻稿2020年1月31日, 新闻稿2019年11月25日, NCT03880474, NCT03883113, NCT03300362, NCT00993083, NCT03277456, NCT02014168, NCT01465035, NCT00942071, 开发者网站:现在被称为Barinthus生物疗法 |
| MVA载体疫苗 | 紧急生物解决方案(美国) | 病毒载体 | 临床前的 | 表达HA、NP、M1、M2、M2e重复序列或串联重复序列(METR)以及具有跨膜区和细胞质环的M2e(m2 etml)的MVA载体疫苗,作为单独或组合递送的单一抗原。 |
| N1·VLP | 佐治亚州立大学(美国)、加拿大卫生部(加拿大) | 病毒样颗粒(VLP) | 临床前的 | VLP含钠从2009 pH1N1旨在诱导交叉反应性NA抑制活性,作为广泛保护性疫苗的组成部分。 |
| N1-I眼镜蛇NA抗原 | 佐治亚大学(美国) | 重组蛋白质 | 临床前的 | COBRA(计算优化的广泛反应性抗原)产生的N1-I NA疫苗设计用于与N1 NA亚型的禽、猪和人流感病毒交叉反应。 |
| NA VLPs | 庆熙大学(韩国南部) | 病毒样颗粒(VLP) | 临床前的 | 禽流感VLP疫苗显示H5血凝素(HA)抗原与或不与禽NA亚型(N1,N6,N8)在不同的组合。 |
| 钠-VLPs | 通武里国王科技大学(泰国) | 病毒样颗粒(VLP) | 临床前的 | 合成并组装成VLPs的重组NA蛋白,旨在诱导抗NA抗体 |
| NA2 VLP | 奥本大学(美国),埃默里-UGA赛尔斯大学(美国) | 病毒样颗粒(VLP) | 临床前的 | 含有来自A/Perth/16/2009 (H3N2)的NA蛋白和来自A/MI/73/2015的基质1 (M1)蛋白的疫苗,采用水包油包水佐剂配制 |
| 奈哈和M2e-哈 | 佐治亚州立大学(美国) | 基于流感病毒 | 临床前的 | 灭活的、具有复制能力的重组流感病毒,其表达具有保守NA表位(NAe)或保守基质蛋白(M2e)的嵌合HA分子或具有插入H3 HA N末端的4M2e表位的嵌合4xM2e-HA融合蛋白;旨在诱导针对保守表位的抗体反应,用于异亚型免疫。 哦2022 (PMID: 35914365), 公园2022 (PMID: 34929590), 公园2021 (PMID: 33603072), 金2020 (PMID: 32882637), 金2017 (PMID: 29107058) |
| 纳米流感 | Novavax(美国),紧急生物解决方案(美国) | 非VLP纳米粒子 | 第三阶段 | 重组体 草地夜蛾(Sf9)昆虫细胞或杆状病毒系统衍生的四价血凝素纳米颗粒流感疫苗(qNIV ),采用基于皂苷的佐剂Matrix-M配制。 Shinde 2022, 伊萨科娃-西瓦克2022 (PMID: 34563276), 阿巴西2021 (PMID: 34751731), 信德2021 (PMID: 33146720), 波特诺夫2020 (PMID: 32098409), Shinde 2018 (PMID: 29897849), 史密斯2017 (PMID: 28844407), NCT04120194, NCT03658629, NCT03293498, 新闻稿2023年5月9日, 新闻稿2021年9月23日, 新闻稿2021年5月10日, 新闻稿2020年10月13日, 新闻稿2020年5月6日, 新闻稿2020年3月24日, 新闻稿2020年2月25日, 新闻稿2020年1月15日, 新闻故事2023年5月9日, 开发者网站:Novavax, 开发者网站:紧急生物解决方案 |
| 纳索瓦克斯 | 高度免疫(美国) | 病毒载体 | 第二相 | 设计用于在鼻上皮细胞中表达流感病毒HA (H1)的复制缺陷型腺病毒血清型5构建体;作为鼻腔喷雾剂给药。 Tasker 2021 (PMID: 33807649), Tasker 2018 (PMCID: PMC6253025), 张2011(:21818346), NCT03760549, NCT03232567, 新闻稿2019年3月20日, 开发者网站 |
| nM2Pr | 韩国生物科学和生物技术研究所(韩国) | 重组蛋白质 | 临床前的 | 基于M2e(nM2e)N末端序列和辅助T细胞表位的线性肽构建体。 |
| NMHC | 中国科学院(中国) | 重组蛋白质 | 临床前的 | 重组蛋白,NMHC,由病毒保守表位和一个超抗原片段组成 |
| NP mRNA | 根特大学(比利时) | 基于核酸的 | 临床前的 | 编码NP的mRNA构建体;经鼻给药;旨在诱导全身性T细胞反应和保护免受异源感染。 |
| NvC-M2ex3 | 马来西亚普特拉大学(马来西亚) | 病毒样颗粒(VLP) | 临床前的 | M2e抗原展示在嵌合蛋白(NvC-M2ex3) VLPs上。 |
| 优化的M2e DNA疫苗 | 中国科学院(中国) | 基于核酸的 | 临床前的 | 以流感病毒M2蛋白(M2e)的N端胞外区为靶抗原;构建了两种优化的M2e DNA疫苗(p-tPA-p3M2e和p-p3M2e),抗原表位密度增加,抗原分泌增强。 |
| OVX836 | 奥西瓦克斯(法国) | 非VLP纳米粒子 | 第二相 | 具有多拷贝全长NP抗原的自组装纳米颗粒;旨在刺激抗体、细胞毒性T细胞和T辅助细胞。 伊萨科娃-西瓦克2023 (PMID: 37517421), 勒鲁-罗尔2023 (PMID: 37517422), withange 2022(PMID:34653245), 勒鲁-罗尔2022 (PMID: 35464450), 德尔坎波2021 (PMID: 34177921), 地平线2020, 德尔坎波2019 (PMID: 30701093), NCT04192500, NCT03594890, NCT05284799, NCT05060887, NCT05569239, NCT05184387, NCT05734040, 新闻稿2023年5月12日, 新闻稿2023年7月28日, 新闻稿2023年6月15日, 新闻稿2023年3月28日, 新闻稿2022年12月21日, 新闻稿2022年9月1日, 新闻稿2022年6月23日, 新闻稿2022年5月25日, 新闻稿2022年5月17日, 新闻稿2022年4月21日, 新闻稿2021年2月12日, 新闻稿2020年9月10日, 新闻稿2020年7月8日, 新闻稿2020年4月9日, 新闻稿2020年1月21日, 新闻稿2019年5月12日, 发展更新2019年12月5日, 开发者网站, 欧盟委员会资助 |
| p22-公顷头VLP | 印第安纳大学(美国) | 病毒样颗粒(VLP) | 临床前的 | 使用SpyTag/SpyCatcher共价附着策略,在多价抗原的高密度展示中,将噬菌体P22 VLPs与HA头结构域的多个拷贝缀合;旨在在快速、可扩展的生产系统中赋予有效的抗体应答(例如,用于爆发情况)。 |
| pABOL配制的saRNA疫苗 | 伦敦帝国学院(英国) | 基于核酸的 | 临床前的 | 可生物还原的阳离子聚合物pABOL,用于递送在初始加强方案中表达来自H1N1或H3N2流感病毒的血凝素(HA)的自扩增RNA (saRNA)疫苗 |
| DNA引物/病毒载体加强策略中的PAPB1M1和NPPB2M2复合免疫原 | 上海公共卫生临床中心(中国) | 基于核酸的 | 临床前的 | 两种新的复合免疫原,其包含克隆到3个连续构建体中的6种甲型流感病毒内部抗原的高度保守的T细胞表位(基于来自约40,000种甲型流感病毒株的M1、M2、NP、PA、PB1和PB2的共有氨基酸序列);DNA引发疫苗,随后用鼻内AdC68病毒载体和肌内天坛痘苗(TTV)病毒载体连续加强。 |
| PapMV-sM2e纳米颗粒 | 拉瓦尔大学(加拿大) | 非VLP纳米粒子 | 临床前的 | 携带M2e和NP抗原的两种类型的PapMV(番木瓜花叶病毒)纳米颗粒;旨在诱导广泛的保护性免疫反应。 |
| pEx 4M2e | 斯洛伐克科学院(斯洛伐克共和国) | 基于核酸的 | 临床前的 | DNA疫苗旨在通过将截短的NS1蛋白的序列插入表达载体pEx (PTriEx-4)中,然后插入4xM2e,从而诱导抗M2e免疫反应。 |
| pH反应性NP疫苗 | 范德比尔特大学(美国) | 非VLP纳米粒子 | 临床前的 | 用于在肺中产生抗原特异性CD8+ TRM细胞的pH响应性聚合物纳米颗粒疫苗;鼻内给药;目的是在肺中诱导快速产生保护性CD8+ TRM细胞。 |
| PIV5-NA | 佐治亚大学(美国) | 病毒载体 | 临床前的 | 表达来自疫情流感(pdmH1N1)病毒或高致病性禽流感(H5N1)病毒的NA (PIV5-NA)的副流感病毒5重组疫苗候选物;旨在引发强有力的交叉反应保护。 |
| PLGA纳米粒子 | 双威大学(马来西亚) | 非VLP纳米粒子 | 临床前的 | 包裹六种保守肽的自佐剂PLGA纳米颗粒 |
| 多价DNA疫苗 | 史泰登血清研究所(丹麦) | 基于核酸的 | 临床前的 | 广泛保护性多价甲型流感DNA疫苗的无针皮内应用,该疫苗编码来自糖基化程度较低的大流行性H1N1 (2009)和H3N2 (1968)病毒株的HA和NA蛋白,以及来自不同大流行性H1N1 (1918)毒株的核蛋白(NP)和基质蛋白(M1和M2)。 吉尔福伊尔2020 (PMID: 33036805), 博格伦2016 (PMID: 27211039), 博格伦2015 (PMID: 25746201) |
| PROTAC | 中国科学院(中国) | 基于流感病毒 | 临床前的 | 蛋白水解靶向嵌合(PROTAC)疫苗技术,使用宿主细胞泛素-蛋白酶体系统有条件地降解流感病毒蛋白,旨在引发针对同源和异源病毒攻击的强大和广泛的体液、粘膜和细胞免疫反应 Si 2022 (PMID: 35788567), 吉尔伯森2022 (PMID: 35869290), 李2022(:36281705) |
| 四价HA mRNA | 绿光生物科学(美国) | 基于核酸的 | 临床前的 | 编码四种季节性流感病毒HA的四价mRNA疫苗 |
| 四价VLP (QVLP) | 苜蓿(加拿大) | 病毒样颗粒(VLP) | 第三阶段 | 烟草植物来源的(Proficia)含HA的四价病毒样颗粒(QVLP);旨在刺激抗体和细胞免疫反应。 阿尔瓦雷斯2022 (PMID: 35750356), 亨丁2022 (PMID: 35410323), 2021病房(PMID: 33581920), 2020病房(PMID: 33065035), 特雷哥宁2020 (PMID: 33065033), Pillet 2019 (PMID: 31166987), 赢得2018年(PMID: 30448064), Pillet 2016 (PMID: 26987887), 新闻稿2023年3月2日, 新闻稿2019年10月1日, 新闻稿06-2018, NCT03739112, NCT03301051, NCT03321968, NCT02831751, NCT02768805, NCT02236052, NCT02233816, NCT01991587, NCT04622592, 开发者网站 |
| rAd-HA2 | 加拿大卫生部(加拿大)、渥太华大学(加拿大) | 病毒载体 | 临床前的 | 重组体腺病毒携带代表所有乙型流感病毒共有序列的合成HA2的疫苗(rAd) |
| rAd-SHA2 | 国家食品和药物控制研究所(中国)、加拿大卫生部(加拿大) | 病毒载体 | 临床前的 | 编码分泌型融合蛋白的重组腺病毒(rAd)构建体(与三聚体形式的鼠CD40L融合的密码子优化的甲型流感病毒HA2亚单位)。 |
| rAd/B-NP | 梨花女子大学(韩国) | 病毒载体 | 临床前的 | 编码乙型流感病毒NP的重组腺病毒(rAd/B-NP);旨在诱导B-NP特异性体液和细胞免疫反应。 孔2023(:37670808), 钟2021 (PMID: 34522441), 金2019 (PMID: 30639307), 李2019 (PMID: 30775351) |
| 用修饰的NS-1和NP重配LAIV | 实验医学研究所(俄罗斯) | 基于流感病毒 | 临床前的 | 重配LAIV表达修饰的NP和NS1基因,旨在诱导强有力的T细胞反应 普罗科彭科2023 (PMID: 36992084), 斯捷潘诺娃2021 (PMID: 33513862), 伊萨科娃-西瓦克2019 (PMID: 31288422), 伊萨科娃-西瓦克2018 (PMID: 29574336), 科伦科夫2018 (PMID: 29929009), 科伦科夫2018 (PMID: 29252117) |
| rM2e-δ板层 | 重庆医科大学(中国) | 重组蛋白质 | 临床前的 | 一种新的含有多个M2e的rM2e-δPly蛋白来源于不同种类的IAV,在大肠杆菌中表达大肠杆菌(大肠杆菌) |
| rMVA-k1-k2 | 联邦医学生物署(俄罗斯) | 病毒载体 | 临床前的 | 基于表位的抗流感疫苗基于改良的安卡拉痘苗(MVA)载体,并使用算法从甲型流感和乙型流感的NP、M1和HA蛋白的保守片段中选择表位 |
| 核糖核酸 | 美国西奈山伊坎医学院 | 重组蛋白质 | 临床前的 | 重组流感病毒N1神经氨酸酶候选疫苗,N1-MPP,辅以TLR9激动剂CpG 1018,旨在有助于开发一种广泛保护性的基于钠的流感病毒候选疫苗。前N1-MPP。 麦克马洪2023 (PMID: 37800945), 莫蒙特2023 (PMID: 37258672), 斯特罗梅尔2022 (PMID: 35869085), 鲁比杜2022 (PMID: 35446141), 谭2022(:34669506), 拉金德兰2021 (PMID: 34451971), 哦2021 (PMID: 34890255), 斯特罗梅尔2021 (PMID: 34809451), 斯特罗梅尔2021 (PMID: 33921722), 麦克马洪2020 (PMID: 32943267), 麦克马洪2019 (PMID: 31113896), 沃博尔德2015 (PMID: 25759506) |
| rNA抗原 | 根特大学(比利时)、赛诺菲巴斯德(美国) | 重组蛋白质 | 临床前的 | 计算设计的共有NA序列形成3种重组NA蛋白。 |
| rNP加BPPcysMPEG | Helmholtz感染研究中心(德国),库约医学和生物实验研究所(阿根廷) | 重组蛋白质 | 临床前的 | 含有用TLR2/6激动剂(BPPcysMPEG佐剂)佐剂化的NP抗原的基于重组蛋白的疫苗 |
| rNP+SLA-SE | 特鲁多研究所(美国) | 重组蛋白质 | 临床前的 | 有佐剂的重组核蛋白构建体;旨在产生针对保守的甲型流感病毒蛋白的细胞和体液免疫。 |
| RSM2eFP孢子疫苗 | 北京微生物与流行病学研究所(中国) | 基于流感病毒 | 临床前的 | 重组流感疫苗使用表达M2e-FP蛋白(RSM2eFP)的枯草芽孢杆菌孢子,通过雾化气管内接种(i.t .)施用。 |
| RVJ-4M2eNP | 中国疾病控制中心(中国) | 病毒载体 | 临床前的 | 表达NP和1或4拷贝M2e的融合抗原的重组痘苗病毒(天坛株)。 |
| rVSV-eδM-tM2e和rVSV-eδM-HM2e | 马尼托巴大学(加拿大) | 病毒载体 | 临床前的 | 基于rVSV的疫苗候选物,其表达流感基质蛋白(M2e)的胞外域和/或保守的血凝素柄区(HA柄),与EboGP的DC靶向域(E1M)融合以产生E1M-tM2e或E1M-HM2e
|
| 甲型流感 | 牛津大学(英国)、墨尔本大学(澳大利亚) | 基于流感病毒 | 临床前的 | 假型流感病毒的非复制形式,通过抑制HA信号序列而失活(S-FLU);旨在通过激活肺中的交叉反应性T细胞来诱导异型保护。 郑2023(:37683004), 鲍威尔2019 (PMID: 30714896), 霍尔茨2018 (PMID: 29703861), 巴兹2015 (PMID: 26489862) |
| sa-mRNA双顺反子疫苗 | CSL·塞奇鲁斯(美国),大角星治疗公司(美国) | 基于核酸的 | 第一相 | 自扩增mRNA双顺反子流感疫苗候选物被开发用于共表达HA和NA 川2023 (PMID: 37794051), 昌2022, NCT06028347, NCT06125691, 新闻稿2022年11月1日, 新闻稿2022年5月10日, 开发者网站:CSL·塞奇鲁斯, 开发者网站:大角星疗法 |
| 萨姆-GM-CSF +萨姆-NP | 葛兰素史克(美国) | 基于核酸的 | 临床前的 | 表达NP的自身扩增mRNA (SAM)疫苗;共同施用SAM-粒细胞-巨噬细胞集落刺激因子(GM-CSF);旨在诱导NP特异性CD8+ T细胞。 马拉纳2019 (PMID: 31227353), Magini 2016 (PMID: 27525409), 布拉佐利2016 (PMID: 26468547) |
| 萨尔纳 | 辉瑞公司(美国) | 基于核酸的 | 第一相 | 肌内递送的自扩增核糖核酸(saRNA)疫苗 |
| 加扰的HA (scrHA) | 威斯康星大学(美国) | 重组蛋白质 | 临床前的 | 一种在HA头部结构域的免疫显性表位具有多种氨基酸的H3 HA疫苗抗原。 |
| 自我扩增mRNA | 明尼苏达大学(美国) | 基于核酸的 | 临床前的 | 编码甲型流感病毒核蛋白的自扩增mRNA疫苗,该疫苗被包裹在修饰的树突状纳米颗粒中。 |
| 展示M2e和HA2的自组装肽 | 俄罗斯科学院(俄罗斯) | 非VLP纳米粒子 | 临床前的 | 展示旨在诱导体液和T细胞反应的M2e和HA柄表位的自组装纳米颗粒 济科娃2023 (PMID: 37764217), 布洛赫纳2023 (PMID: 37299207), 济科娃2022 (PMID: 34583058), 济科娃2020 (PMID: 33036278) |
| 自组装蛋白质纳米笼 | 佐治亚理工学院(美国) | 非VLP纳米粒子 | 临床前的 | 一种广泛交叉反应的流感疫苗,通过功能化自组装蛋白质纳米笼(SAPNs ),在外表面上有多个血凝素柄拷贝,在内表面上有基质蛋白2胞外域 |
| 自组装蛋白质纳米颗粒 | 斯克里普斯研究所(美国) | 非VLP纳米粒子 | 临床前的 | 呈现基质蛋白2 (M2e)保守胞外结构域的单组分自组装蛋白纳米颗粒(SApNPs) |
| 单ADCC激活肽 | 香港大学(中国香港特别行政区) | 重组蛋白质 | 临床前的 | 基于HA1和HA2区ADCC诱导肽的新构建体。 |
| 斯皮塔格北VLP | 坦佩雷大学(芬兰) | 病毒样颗粒(VLP) | 临床前的 | 使用基于诺罗VLP的模块化疫苗平台,融合了高度保守的抗原、M2e蛋白的胞外域和HA的最小化茎片段。 兰皮宁2023 (PMID: 37424789), Heinimaki 2022 (PMID: 34894525), 兰皮宁2021 (PMID: 33468139) |
| 稳定的无头HA茎纳米粒子 | 美国国家过敏和传染病研究所 | 非VLP纳米粒子 | 第一相 | 自组装纳米粒子上稳定的无头HA干细胞三聚体:旨在广泛刺激针对新型病毒的保护性免疫。 安德鲁斯2023 (PMID: 37075126), Widge 2023 (PMID: 37075129), 莫因2022 (PMID: 36356572), 达里卡尔2021 (PMID: 33658355), 科比特2019 (PMID: 30808695), 亚辛2015 (PMID: 26301691), 新闻稿2023年4月20日, 新闻稿2021年3月3日, NIH新闻发布会2019, NCT03814720, NCT04579250, NIH资助, 开发者网站:NIAID |
| 合成多组分纳米疫苗 | 蒙特利尔魁北克大学(加拿大) | 非VLP纳米粒子 | 临床前的 | 多组分纳米丝,由交叉β-折叠纳米丝组成,暴露流感病毒和TLR7激动剂IMQ的M2e表位,旨在诱导保护性M2e特异性免疫反应。 |
| T4-M2e VLP | 华中农业大学(中国) | 病毒样颗粒(VLP) | 临床前的 | 展示来自人、猪和禽流感的M2e的三个串联拷贝的T4 VLP构建体;旨在诱导体液和细胞免疫反应。 |
| ta-RNA,sa-RNA | BioNTech(德国),约翰内斯·古腾堡大学美因茨(德国) | 基于核酸的 | 临床前的 | RNA疫苗方法基于使用编码HA抗原的反式扩增RNA (taRNA)或自我扩增RNA(saRNA)的新型分裂载体系统。 佩尔科维奇2023 (PMID: 36694464), 贝思特2020 (PMID: 31624015), 沃格尔2018 (PMID: 29275847) |
| VLP坦迪弗卢 | 伊克尔(英国) | 病毒样颗粒(VLP) | 临床前的 | 基于HepB串联核心平台上呈递的多种保守抗原的VLP构建体:在酵母中表达的HepB核心蛋白+ HA茎(H1和H3) + 3xM2e(来自3个菌株);一个VLP中含有4个保守的IAV抗原;旨在刺激交叉反应和保护性多克隆抗体。 拉米雷斯2018 (PMID: 29306508), 豪克2017 (PMID: 29587397), 哈萨克斯坦2017 (PMID: 29126399) |
| TAT-NP | 上海生物制品研究所(中国) | 重组蛋白质 | 临床前的 | 基于保守蛋白NP和蛋白转导结构域TAT的重组蛋白构建体;旨在诱导交叉保护性细胞和体液免疫反应。 |
| 三聚体HA-Fc | 马里兰大学(美国) | 重组蛋白质 | 临床前的 | 单体IgG Fc与流感病毒血凝素(HA) Ag融合,具有三聚化结构域 |
| TX98-129 | 南达科他大学(美国)、伊利诺伊大学厄巴纳-香槟分校(美国) | 基于流感病毒 | 临床前的 | 表达嵌合HA (HA-129)的基于重组流感病毒的疫苗,所述嵌合HA(HA-129)衍生自具有人畜共患病传播史的四种遗传上不同的甲型猪流感病毒(H1N1)的HAs。嵌合HA是通过分子育种(基因改组)技术从四个原始亲本HA中获得的。旨在诱导广泛的保护性免疫来对抗遗传上不同的HAs。 |
| VLP鸡尾酒 | 美国国家过敏和传染病研究所 | 病毒样颗粒(VLP) | 临床前的 | 分别显示H1、H3、H5和H7的VLP混合物代表第1组和第2组病毒;旨在抵御正在传播的新型甲型流感病毒。 施瓦茨曼2015 (PMID: 26199334), 舒尔茨樱桃2015 (PMID: 26443464), 新闻稿2015年7月21日 |
| VLP疫苗 | TechnoVax(美国) | 病毒样颗粒(VLP) | 临床前的 | 显示重塑HAs的替代构象的VLP结构显示保守的亚优势抗原位点;旨在促进抗体反应以中和广谱流感病毒。 马特索伊2007 (PMID: 17931114), 新闻稿2018年9月12日, 授权摘要1, 授权摘要2, 开发者网站:TechnoVax |
| VRC H1ssF_3928 mRNA-LNP | 美国国家过敏和传染病研究所 | 基于核酸的 | 第一相 | 与幽门螺杆菌铁蛋白共价融合的表达流感H1稳定干(H1ss)的mRNA |
| VT-105 | Versatope Therapeutics(美国)、Intravacc(荷兰) | 非VLP纳米粒子 | 临床前的 | 结合了展示在单个纳米大小的rOMV(重组外膜囊泡)上的不同流感毒株变体;VT-105基于M2蛋白;靶向病毒感染的细胞,以参与针对所有流感毒株的细胞介导的免疫反应。 Weyant 2023 (PMID: 36709333), 沃特金斯2017 (PMID: 28215994), 沃特金斯2017 (PMID: 28866291), 拉帕佐2016 (PMID: 26827663), 新闻稿2020年6月8日, 新闻稿2020年3月18日, 新闻故事2017年9月21日, 开发者网站:Versatope Therapeutics |
| VXA-A1.1口服片剂 | 瓦克萨特(美国) | 病毒载体 | 第二相 | 表达HA的复制缺陷型腺病毒5型载体构建体;包括作为佐剂的新型toll样受体3 (TLR3配体)激动剂;口服片剂,用于在回肠释放病毒,有可能刺激细胞和粘膜免疫以及血清抗体。 博朗2023 (PMID: 37562075), Flitter 2022 (PMID: 35455342), McIlwain 2021 (PMID: 34784508), 鲁登科2020 (PMID: 31978351), 利博维茨2020 (PMID: 31978354), Kolhatkar 2018 (PMCID: PMC6253129), 斯考兰2016 (PMID: 27071663), 金2016 (PMID: 27881837), 利博维茨2015 (PMID: 26333337), 彼得斯2013 (PMID: 23357198), NCT02918006, NCT03121339, NCT01335347, 新闻稿2020年1月22日, 新闻稿2019年7月9日, 开发者网站 |
| 惠氏/白介素-15/5流感 | 香港大学(中国香港特别行政区) | 病毒载体 | 临床前的 | 活的、可复制的牛痘Wyeth骨架,其携带来源于H5N1病毒(NP、HA NA、M1和M2)的5种全长流感蛋白和作为分子佐剂的IL-15;旨在引发强大的CD4+和CD8+ T细胞反应。 公牛2023 (PMID: 36401824), 公牛2022 (PMID: 35385318), 2018年瓦尔肯堡冬奥会(PMID: 29887326), 瓦尔肯堡2014 (PMID: 24706798) |
| α-1,3-GT | 香港大学(中国香港特别行政区) | 基于流感病毒 | 临床前的 | 表达半乳糖-α-1,3-半乳糖(α-Gal)表位的LAIV;旨在增强抗原更新,导致吞噬作用、NK细胞介导的细胞杀伤和ADCC反应以及针对1型和2型流感病毒的交叉反应保护;鼻内给药。 |
| α-DEC-205: M2e结合物 | 莫雷洛斯州自治大学(墨西哥) | 重组蛋白质 | 临床前的 | M2e肽与DEC-205(一种存在于树突细胞上的C型凝集素)特异性单克隆抗体化学交联,以诱导异亚型保护。 |
| NS1病毒 | 美国西奈山伊坎医学院 | 基于流感病毒 | 临床前的 | 缺乏NS1基因的减毒活甲型流感病毒鼻内给药,旨在诱导先天性抗病毒反应 |
通用流感疫苗技术前景 |CIDRAP公司 (umn.edu)
Hits: 82