
任何能够引起宿主生物疾病的微生物都被称为病原体 。本文仅限于人类微生物病原体,尽管动植物病原体在自然界也广泛存在。当致病微生物 ( 细菌 、 病毒或原生动物寄生虫 ) 感染人体时,宿主的先天性和适应性免疫系统与病原体的各种毒力机制和因子之间便会展开一场较量。这场较量的结果决定了宿主能否存活和康复,以及康复程度如何。完全康复需要宿主恢复生理(和免疫)稳态 ,而恢复所需的时间取决于感染的性质和严重程度,以及是否采取了任何预防或治疗措施。许多病原体还会运用多种免疫逃逸策略来入侵和定植宿主细胞,并可能成功利用宿主细胞进入靶组织。
本文介绍了一些毒力策略和免疫逃逸策略,其中免疫逃逸策略可能包括:(1) 躲避免疫系统(例如,藏身于细胞内);(2) 干扰免疫系统功能 (例如,阻断信号);(3) 破坏免疫系统成分 (例如,将微生物抗原呈递给免疫效应细胞以启动宿主免疫反应的结构)。 毒力通常涉及利用各种机制破坏宿主细胞或导致宿主细胞功能障碍。宿主也可能采取反制策略来应对这些攻击。关于特定微生物所采用的各种机制的更详细描述,请参阅其他“简明扼要”页面。
水痘病毒(Varicella zoster )和疱疹病毒科 ( 单纯疱疹病毒、水痘-带状疱疹病毒、巨细胞病毒等 )等病毒可以潜伏在神经元和非神经元细胞中,逃避免疫系统的攻击,并在宿主抵抗力下降时以致病形式出现。伯氏疏螺旋体 (Borrelia burgdorferi) 和类鼻疽伯克霍尔德菌 (Burkholderia pseudomallei )(分别引起莱姆病和类鼻疽 )等细菌也存在类似情况,有报道称,感染症状在初次感染后数月至数年( 伯氏疏螺旋体)甚至长达 60 年( 类鼻疽伯克霍尔德菌 )内复发。在免疫干扰方面,病原体可能会采取更明显的策略,例如利什曼原虫属(Leishmania spp.)的寄生虫。能够选择性地抑制宿主体内促炎细胞因子白细胞介素 12 ( IL12p40 ) 的转录,从而抑制宿主的免疫反应。
为了维持其毒力机制,许多细菌能够通过合成铁结合载体(铁载体)来螯合哺乳动物宿主体内的游离铁 。铁是宿主和微生物代谢的必需成分。因此,为了保护自身免受此类毒力机制的侵害,宿主细胞会合成铁载体(铁载体)来对抗细菌 ,这些受体能够竞争性地结合铁。哺乳动物宿主细胞还进化出了一系列识别微生物或微生物因子的模式识别受体 ,例如 Toll 样受体 (TLR),这些受体与微生物或微生物因子结合后,会触发细胞内信号级联反应,从而产生抗菌作用。
虽然许多细菌病原体本质上是细胞内寄生的,但也有一些病原体无需侵入宿主细胞,而是利用各种分泌过程将毒素和其他毒力因子递送至宿主细胞内。例如,大肠杆菌、福氏志贺氏菌、鼠疫耶尔森氏菌和沙眼衣原体等细菌已经进化出形成空心突起(即所谓的转位子 )的能力,该突起在接触宿主细胞后可将抗宿主因子递送至宿主细胞内,通常会导致宿主细胞凋亡(即所谓的 III 型分泌 )。这些细菌分别引起食物中毒、痢疾、 鼠疫和泌尿生殖道感染等多种疾病(见图 1 )。然而,一些细菌,例如土拉弗朗西斯菌 ( 土拉菌病的病原体)和伯克霍尔德氏菌属,则不具备这种能力。 (引起类鼻疽或鼻疽的 )具有多种分泌过程,通过这些过程将毒力因子输送到宿主细胞中。
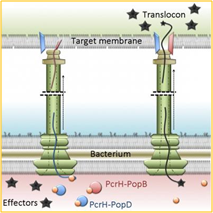
另一种严重的人类病原体, 炭疽杆菌( 引起炭疽病的病原体 ), 具有完善的毒力机制,涉及三种蛋白质的分泌。其中一种蛋白质, 保护性抗原 (PA),可与宿主细胞受体结合,从而促进致死因子 (LF)或水肿因子 (EF)的进入。PA-LF 或 PA-EF 复合物经网格蛋白介导的内吞作用进入早期内体 ,随后通过微管运输,经由囊泡进入酸化的晚期核周内体 。在这些条件下,LF 被释放到细胞质中,而 EF 则仍然结合在晚期内体的核周膜上。在细胞质中,LF 切割并灭活丝裂原活化蛋白激酶激酶 (MAPKK),从而破坏细胞核内的磷酸化和转录,最终阻止蛋白质合成并导致细胞死亡。 EF 是一种钙和钙调蛋白依赖性腺苷酸环化酶,它能迅速增加细胞核周围的 cAMP 水平,导致细胞、组织乃至最终的器官水肿。LF 和 EF 均能抑制促炎细胞因子的分泌 ,并通过下调血管钙黏蛋白 (一种在细胞间黏附中起重要作用的蛋白)来削弱血管内皮屏障;这些作用共同导致了全身性炭疽典型的血管渗漏。
另一方面,病毒病原体确实需要侵入宿主细胞才能完成其复制周期。近年来出现的最严重的病原体之一—— 人类免疫缺陷病毒 (HIV)会劫持 CD4+ T 细胞 ,从而削弱宿主产生强效细胞介导免疫(CMI)反应的能力。HIV 的免疫逃逸策略影响范围极广,迄今为止,这阻碍了有效疫苗的研发进展。首先,HIV 会通过免疫豁免部位(例如阴道或直肠)进入未感染者的体内,这些部位淋巴组织稀少;随后,病毒利用辅助受体 ( CCR5/CXCR4 )与 CD4+宿主细胞结合。其结果是 CD4+ T 细胞逐渐减少,宿主细胞介导免疫(CMI)功能减弱,宿主对其他感染(如细菌性肺炎)或肿瘤(如卡波西肉瘤)的易感性增加 。HIV 感染病例目前通过抗逆转录病毒药物进行控制,同时疫苗候选物正在研发和评估中。 天花病毒 (引起天花的病原体)也进化出了逃避宿主免疫的策略,它分泌一种抑制补体酶活化的蛋白质,以及另一种趋化因子结合蛋白 II 型 ,后者能够阻断感染部位免疫细胞和炎症所需的信号。病原体还可以通过干扰表面 MHC I 类蛋白的表达来逃避宿主的抗原加工和呈递系统,但这反过来又会增加它们对自然杀伤(NK)细胞活性的敏感性。 然而,疱疹病毒、乳头瘤病毒、逆转录病毒、痘病毒和黄病毒家族的成员也进化出了逃避自然杀伤细胞攻击的机制,并通过抑制宿主细胞凋亡来促进自身在宿主细胞内的存活。最后,一些病原体经常改变其表面抗原 ,流感病毒(正粘病毒)就是一个典型的例子。
通过所有这些毒力机制和免疫逃逸技术,病原体已经进化出在宿主体内生存的策略。当然,病原体与宿主的相互作用是一个动态过程,病原体取得的任何进展都会被宿主所抵消。因此,宿主体内细胞毒性 T 细胞(Tc)的扩增会导致病原体发生逃逸突变 ,从而避免成为 Tc 的靶标。因此,病原体将不断进化和出现,而可以说,最成功的病原体是那些利用宿主而不杀死宿主的病原体。
© 本作品的版权归作者所有。
Host − Pathogen interactions and immune evasion | British Society for Immunology
Hits: 24
老师晚上好。过来打扰了,事情是这样。我买了两瓶饮料。当时这两瓶饮料恰好有一条狗舔了外面,就是那个塑料瓶。我的疑问就是他不会穿透塑料瓶进入饮料里吧,我还把这事给忘了,回来就喝了,2月份打的加强针。这种情况需要担心吗?感谢博士,辛苦您了。
安全