
Contents
Concurrent Clade I and Clade II Monkeypox Virus Circulation, Cameroon, 1979–2022
摘要
在1979-2022年期间,在国家监测网络确定的137例疑似mpox病例中,喀麦隆记录了32例实验室确认的mpox病例。最高的阳性率出现在2022年,表明mpox可能在喀麦隆重新出现。进化枝I (n = 12)和进化枝II (n = 18)猴痘病毒(MPXV)均有报道,这是喀麦隆mpox的一个独特特征。2.2%的总病死率与clade II相关。我们发现mpox只发生在该国南部的森林地区,MPXV的系统地理结构揭示了并行循环进化枝之间明显的地理分离。进化枝I起源于中非邻近mpox流行国家的东部地区;进化枝II在靠近西非的西部地区流行。我们的发现表明,MPXV在30年后重新出现,可能来自喀麦隆东部和西部雨林生态系统特有的不同病毒宿主。
猴痘病毒是一种新出现的人畜共患病病毒正痘病毒属导致人类mpox,一种类似于根除天花的疾病(1)。自从1958年在一只猴子身上发现后(2)和一个人类在1970年(3),MPXV相关疫情主要发生在中非和西非国家的农村雨林(4–6)。
Mpox的特征是伴有腺病和斑丘疹的流感样综合征,典型地出现在手掌和脚底(4,7)。对于感染者,提供支持性护理和抗病毒治疗,包括西多福韦和tecovirimat(4)。天花疫苗的交叉免疫和新一代天花疫苗同样提供了一些保护(8–10)。然而,在20世纪80年代早期停止天花疫苗接种后,群体免疫力逐渐下降,使得mpox重新出现,这在过去30年中非洲病例数量的增加中得到强调(4,8,11–13)。自2022年初以来,病例数激增,截至2022年12月28日,非洲报告了约1,215例mpox确诊病例和219例死亡(14)。在2022年4月之前,西半球的mpox病例通常是通过接触外来宠物贸易和国际旅行(15–20)。自那以后,MPXV相关的疾病在世界范围内爆发,影响了非洲以外的100多个国家(4,21)并成为一个全球性的公共卫生问题。
原发性MPXV传播可通过直接接触受感染动物的体液或皮肤病变发生,或通过受污染的污染物间接发生。与受感染的人或受感染的呼吸道飞沫的类似接触也可能导致人与人之间的二次传播,这是2022年全球爆发的主要传播方式(4,22)。从历史上看,原发性人畜共患病传播更为常见,主要涉及猎人、屠夫和野味加工者等高危人群;继发性传播是罕见的,但医院和家庭传播已被描述(3,13,23–25)。
系统发育研究报告了2个不同的MPXV分支:分支I,在中非流行,分支II,西非特有(5,6,26–28)。然而,喀麦隆是一个例外,两个分支同时在该国流通(6,29)。进化枝I进一步细分为谱系1-5,进化枝II进一步细分为子类IIa和IIb进化枝IIb对2022年开始的多国疫情负责(27,28,30)。全球范围内,MPXV致死率从1%到10%不等,已知进化枝I的死亡率高于进化枝II(4,24,25)。MPXV动物宿主尚未确定,但该病毒可感染多种哺乳动物绳蜥松鼠和洛氏石斑鱼小鼠被认为是最有可能的MPXV储库(31–33)。
在喀麦隆,2022年爆发前仅记录了4例确诊的mpox病例,1979年、1980年、1989年和30年后的2018年各1例(29,34–36)。根据公共卫生报告,该国可能发生了更多病例并且没有记录在案,特别是在2018-2021年期间,尤其是在2022年,在此期间发生了前所未有的mpox疫情,并出现了反复出现的病例群(37)。然而,这些感染是与来自邻国的输入有关,还是来自本地原发或继发传播,仍不清楚(29)。总的来说,关于喀麦隆MPXV发生和传播动力学的流行病学特征的数据很少。我们调查了喀麦隆MPXV相关疾病暴发的临床、流行病学和分子特征。
样本位置
喀麦隆位于非洲中部,划分为10个行政区。喀麦隆因其多样的农业生态背景而被称为非洲的缩影:远北、北部和阿达马瓦地区的草原和稀树草原;沿海和西南地区的海岸带;西北和西部地区的山地高原;以及中部、南部、西南部和东部地区的雨林(38)。喀麦隆有三大热带森林:横跨东部、南部和中部地区的刚果盆地森林;西部和阿达马瓦地区的几内亚潮湿森林;和西南地区的喀麦隆高地森林。一些水道穿过这些森林,包括喀麦隆最大的河流萨纳加河(33–40;J. Thia,坎特伯雷大学硕士论文,2014年,https://www . researchgate . net/publication/272494772尼日利亚山区树木的困境外部连接)。
试样收集
我们将疑似病例定义为>1临床体征或症状,包括头痛、乏力、腺病、与发热相关的肌痛,或逐渐发展为蔓延至身体其他部位的皮疹,包括脚底和手掌。我们将疑似病例定义为没有病毒学证实的临床表现,但与另一个疑似或确诊病例有流行病学联系。确诊病例是实验室确诊的任何MPXV病例。
我们记录了1979-2022年间所有疑似病例的流行病学数据,包括人口统计学和临床信息。我们从同意接受检测的患者身上采集了5毫升血液样本、囊泡拭子、表皮样本或样本组合。我们在三重包装系统下将样品运送到喀麦隆巴斯德中心(CPC ),这是喀麦隆mpox诊断的国家参考实验室。我们排除了无法采集样本的患者。
MPXV感染的实验室确认
在CPC,样本在生物安全3级实验室中接收、处理和灭活。我们通过使用QIAamp DNA微型试剂盒(QIAGEN,https://www.qiagen.com外部连接)按照厂家的说明。如前所述,我们通过通用实时PCR Taqman试验检测了纯化的MPXV DNA(41)。对于显示周期阈值(Ct)值< 37的阳性样品,我们通过使用特异性靶向MPXV分支I和II(41)。
我们进一步扩增了来自2022年爆发的具有Ct值的8个阳性样本的子集<20通过使用靶向MPXV A型包涵体的一部分的PCR(天线调谐电感)基因,根据先前描述的方案(42)。我们使用1%绿色染色的琼脂糖凝胶来显示产生的扩增子,我们将其送至商业服务提供商Inqaba Biotechnical Industries(南非比勒陀利亚)进行Sanger测序。
系统发育分析
我们用CLC Main Workbench软件(QIAGEN)组装新测定的序列并校正。我们通过使用MAFFT第7版(https://mafft.cbrc.jp外部连接)和来自GenBank的56个MPXV参考基因组的扩展数据集(附录表1、2)。我们向软件集成模型查找程序(IQ-TREE,http://www.iqtree.org外部连接)选择基于贝叶斯和赤池信息准则的最佳进化模型。我们使用IQ-TREE版本1.6.12(http://www.iqtree.org外部连接)来推断MPXV上的最大似然进化树天线调谐电感基于Hasegawa-Kishino-Yano加氨基酸替代模型的序列,应用1,000个引导复制。我们向GenBank提交了新确定的序列(登录号or 038717–24)(附录表2)。
统计分析和绘图
为了提供喀麦隆mpox流行病学的完整情况,我们将4例先前记录的喀麦隆mpox病例以及从文献和卫生部档案中收集的可用信息添加到我们的数据集中(29,34–36)。我们通过使用分类变量的频率总结了社会人口统计学和临床特征;我们使用中位数和四分位间距(IQR)作为定量变量。我们使用Pearson χ比较PCR确认的病例和未确认的疑似病例2或分类变量的Fisher精确检验和数量变量的Wilcoxon检验。我们使用单变量逻辑回归来确定与MPXV感染相关的因素,并估计粗优势比(ors)和95% ci。我们无法推断多变量分析模型,这些模型无法收敛是因为丢失了太多数据(表1, 2)。我们认为p<0.05具有统计学显著性,p<0.07具有边缘显著性。我们在R版本4.1(统计计算的R基础,https://www.r-project.org外部连接)。我们用的是量子GIS 3 . 30 . 1版(QGIS,https://qgis.org外部连接)按卫生区域和地理数据分析和绘制mpox病例。
伦理学
样本收集和实验室分析是在喀麦隆国家监测方案的框架内进行的。根据该计划,在我们提供了详细的信息并解释了取样目的后,我们获得了所有疑似mpox患者的书面或口头知情同意。我们获得了15岁以下儿童的父母或公认监护人的知情同意。
在喀麦隆的mpox监测系统中,1979-2022年间,我们确定了137例疑似mpox病例,其中男性74例(54.41%),女性62例(45.59%);1例性别数据缺失(表1)。病例患者的中位年龄为11岁(范围2周到75岁;IQR 4-27岁);近一半(48.18%)是<10岁(表1)。
分子诊断结果
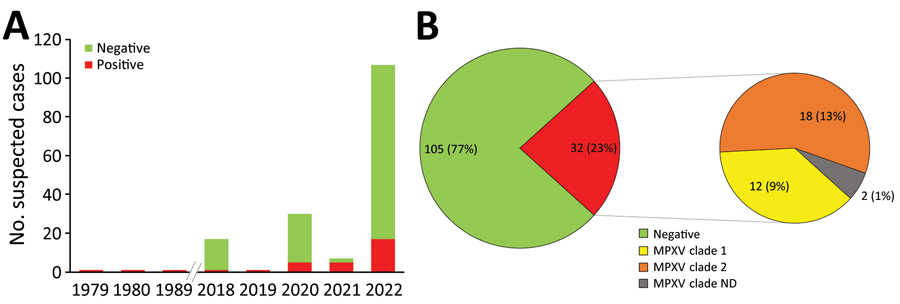
Mpox病毒通用PCR显示,在喀麦隆1979年至2022年期间检测的137例患者中,有32例(23.36%)实验室确认的mpox病例(表1; 图1,面板A;附录表3)。2018年以前,仅有3例散发病例被确认为人感染MPXV。在没有报告mpox病例的30年间隔后,监测系统在2018-2022年期间不断发现新的mpox病例。疑似病例中,2018年仅发现1例,2019年发现1例。2020年和2021年,每年记录5例实验室确诊病例。在2022年期间,mpox病例急剧增加到84例疑似病例中的17例确诊病例(图1,面板A;附录表3)。
在137例疑似病例中,实时PCR结果的基因分型鉴定出12例(9%)患者感染了MPXV分支I,18例(13%)患者感染了MPXV分支II;2例(1%)历史确诊病例缺乏分支判定结果(表1; 图1,面板B;附录表2、3)。在所有实验室确认的病例中,仅记录了1例死亡,患者感染了MPXV进化枝II。卫生部调查记录显示,在具有典型mpox临床表现的人群中,有另外2例患者死亡,这些患者在流行病学上与2例确诊病例-感染了clade II MPXV菌株的患者相关。但是,死前没有采集到标本;因此,我们考虑了那些可能的情况。包括可能病例在内,喀麦隆确诊和疑似病例的总病死率(CFR)为2.2% (3/139),所有死亡均与病毒进化枝II相关。
确诊Mpox病例的流行病学和临床特征
单变量分析显示,按性别划分的感染可能性的增加没有统计学显著差异:21/74 (28.38%)男性和11/62 (17.74%)女性已确诊MPXV感染(表1)。MPXV-确诊病例-患者的中位年龄为21.5岁(范围为2周-52岁;IQR 8.5岁至32.25岁)。MPXV感染在成人中更为普遍>20周岁以下;总的来说,35.56%确诊为MPXV感染,而在年轻的MPXV确诊病例患者中为17.78%(p = 0.025)。然而,我们发现1980年以前出生的成年人与其他人群相比没有统计学上的显著差异(p = 0.092)。需要更大的数据集来证实观察到的趋势。
MPXV感染主要与从事农业的职业活动有关(OR 3.83,95% CI 0.44–33.11)(表1)。同样,在卫生工作者中也发现了潜在的院内传播(OR 3.07,95% CI 0.84–11.17)。其他活动,包括教学、交易或驾驶,当综合考虑时,似乎也是二级MPXV传播的潜在风险活动(OR 2.13,95% CI 0.52–8.77)。然而,在29.82%的MPXV确诊病例中,我们发现与继发性传播无关,这些病例报告了过去与具有mpox样临床症状的人的接触(表1)。因为mpox是典型的动物源性疾病,我们也评估了动物暴露的前因。我们没有观察到与不明动物接触相关,但观察到报告与野生动物接触(OR 2.86,95% CI 0.53–15.47)的确诊病例(6/13 [46.15%])比报告与家畜接触或没有与动物接触的人具有更高的风险(表1)。在接触的野生动物中,研究参与者经常提到松鼠、蝙蝠、毛虫、穿山甲、老鼠、豪猪和猴子。
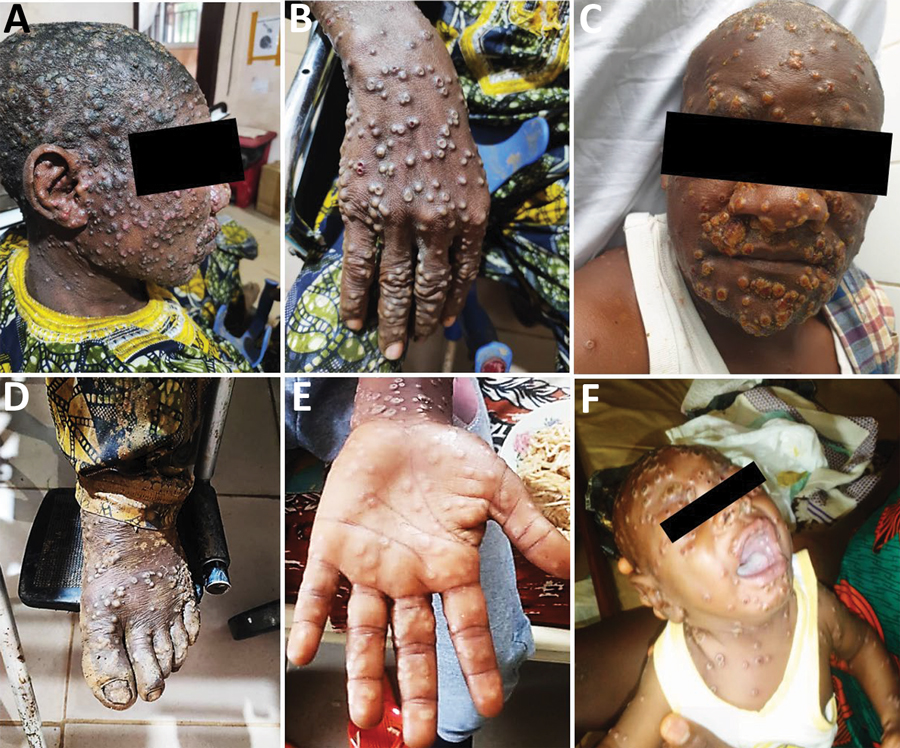
根据要求出现皮疹的病例定义标准,几乎所有(124/137 [90.5%])疑似MPXV病例均出现活动性皮损(表2; 图2; 附录表3)。然而,与MPXV阴性患者相比,我们没有观察到MPXV确诊患者在病变进展、深度、大小或分期方面的具体差异(表2)。在手掌和脚底有病变的确诊病例中,斑丘疹性病变更为普遍(图2)。临床数据确定了咳嗽(OR 2.3,95% CI 0.95–5.59)、发冷或出汗(OR 3.8,95% CI 1.52–9.48)、淋巴结病(OR 3.53,95% CI 1.38–9.00)、吞咽时咽痛(OR 10,95% CI 3.69–27.12)、口腔溃疡(OR 8.38,95% CI 2.8–25.09)和全身疲劳(OR 3.06表2; 图2)。在所有疑似病例患者中,约26%在皮疹出现前报告有发热症状的患者被确诊为MPXV感染,但我们没有发现有无发热的确诊病例之间存在差异。此外,MPXV确诊或阴性的病例在头痛(表2)。我们注意到,与感染进化枝II的病例相比,感染进化枝I的病例在临床严重程度上几乎没有差异(附录表4)。暴露途径也是如此;我们没有发现人畜共患病或人与人之间的传播与特定的感染病毒分支(附录表4)。然而,由于大量数据丢失(表1, 2)我们无法进行多变量分析。因此,很难对喀麦隆mpox感染的流行病学和临床特征做出结论性解释。
地理和系统发育分析
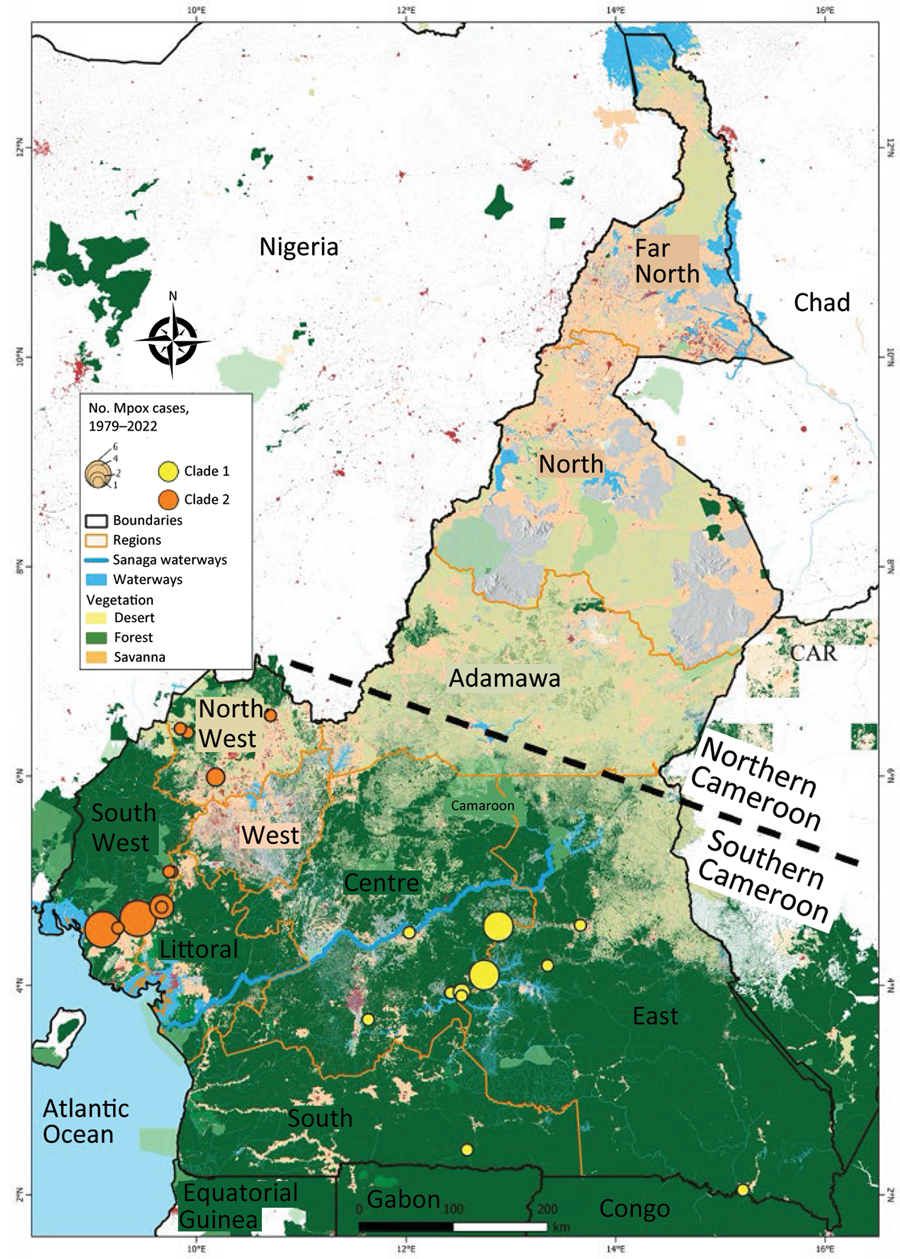 图3.1979-2022年喀麦隆猴痘病毒循环并发分支I和分支II研究中确诊mpox病例和分支的地理分布。在mpox监测系统框架内,共报告了137例疑似mpox病例,其中37例经PCR证实为猴痘病毒感染。进化枝I (12例)和进化枝II (18例)病毒株被确定在该国流行。我们注意到,中部、南部和东部地区与西北部和西南部地区之间存在明显的地理隔离,前者只报道了进化枝I(黄点),后者只发现了进化枝II(橙点)。每个点的大小与地图上确诊病例的数量成正比。该地图是使用Quantum GIS 3 . 30 . 1版(QGIS,https://qgis.org).中非共和国。
图3.1979-2022年喀麦隆猴痘病毒循环并发分支I和分支II研究中确诊mpox病例和分支的地理分布。在mpox监测系统框架内,共报告了137例疑似mpox病例,其中37例经PCR证实为猴痘病毒感染。进化枝I (12例)和进化枝II (18例)病毒株被确定在该国流行。我们注意到,中部、南部和东部地区与西北部和西南部地区之间存在明显的地理隔离,前者只报道了进化枝I(黄点),后者只发现了进化枝II(橙点)。每个点的大小与地图上确诊病例的数量成正比。该地图是使用Quantum GIS 3 . 30 . 1版(QGIS,https://qgis.org).中非共和国。
报告的疑似mpox病例来自喀麦隆的8个行政区(表1; 图3)。大多数(97.08%)疑似病例来自该国南部,那里也是所有确诊病例的发源地。特别是,沿海地区确诊1例(3.13%),南部地区确诊1例(3.13%),东部地区确诊3例(9.38%),西北部地区确诊6例(18.75%),西南部确诊10例(31.25%),中部地区确诊11例(34.88%)(表1; 图3; 附录表3)。值得注意的是,在沿海地区确诊的一例独特病例最初来自西南部,在沿海地区寻求医疗保健。实时PCR的基因分型显示,在来自中部、南部和东部地区的患者中证实了所有分支I MPXV感染;除了1例以外,所有的进化枝II MPXV样本都是从沿海、西北和西南地区的患者身上获得的。事实上,在中部地区发现的进化枝II MPXV是来自西北地区的国内流离失所者(表1; 附录表3)。mpox病例的分布表明喀麦隆2个病毒分支的地理隔离。这些发现表明,在喀麦隆南部,MPXV基因型具有很强的地理关联性,MPXV分支II与该国西部相关,分支I与该国东部相关。
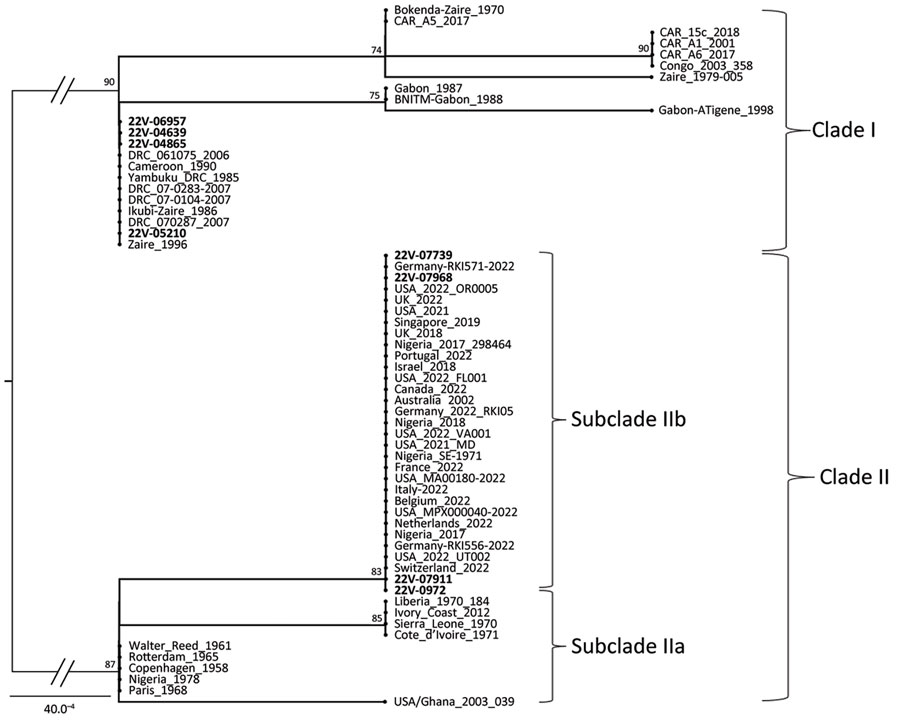
我们得到了部分MPXV天线调谐电感喀麦隆4个地区8例mpox确诊病例的基因序列。我们从西北地区(CPC代码22V-0972)、西南地区(CPC代码22V-07739、22V-07911、22V-07968)、中部地区(CPC代码22V-05210、22V-04865、22V-4639)和南部地区(CPC代码22V-6957)收集的样品中获得了新确定的序列。942个nt共有序列的最大似然种系发生分析,包括参考序列(附录表1-3)显示,来自喀麦隆的8个MPXV基因组分成进化枝I和进化枝II。正如我们报道的MPXV分离株的地理关联所预期的,来自中部和南部地区的MPXV分支I与先前报道的来自中非国家的参考对应物可靠地分组,来自西北和西南地区的分支II序列与来自西非的菌株一致地分组(图4)。来自喀麦隆的进化枝II菌株可靠地聚集在亚分类IIb内,具有83%的引导支持(图4)。总之,基因型和系统发育分析证实了在喀麦隆MPXV分支I和II的同时传播,具有显著的地理隔离。
我们研究了喀麦隆44年间(1979-2022)MPXV感染的临床、流行病学和分子模式,作为该国mpox监测的一部分。在1979-2022年期间,共有137人被怀疑患有mpox,32人被证实感染了MPXV。三人死亡(病死率2.2%),死亡与MPXV分支II相关。该CRF远低于先前MPXV分支I研究中报道的CFR为7%-10%(13,43)。总体而言,在感染clade II的患者中,CFR较低,包括在2022年全球疫情背景下(4,25)。我们无法收集关于病例患者潜在潜在疾病的信息,以确定免疫妥协状况是否导致死亡,这可能会恶化临床疾病表现,正如其他人所强调的那样(44)。此外,与clade I相关的死亡病例可能逃脱了喀麦隆的国家监测系统,该系统是新的,仍在改进中。
我们发现,原发性人畜共患病和继发性人与人之间的MPXV传播都发生在喀麦隆,包括影响卫生工作者的医院传播。我们的结果与描述二级传播链的报告一致,包括在流行国家通过贸易、运输、狩猎和医疗保健进行的家庭内传播和职业传播(24,43,45,46)。这项研究强调了流行国家中常见的MPXV感染途径、种间传播以及野生动物被认为是该病毒的宿主(31,32,47)。区分一次传播和二次传播是困难的,因为两者都可能发生。需要额外的数据和进一步的调查,以清楚地了解喀麦隆MPXV传播的潜在驱动因素。
这项研究的局限性是我们无法进行更精确的分析来确定喀麦隆mpox流行病学的独立特征。因为当前的监视系统仍然是手写的,并且表格经常不完整地填写,所以数据丢失,这在基于纸张的数据收集系统中是常见的(48)。
自1979年以来,喀麦隆10个行政区中的6个发生了MPXV感染:中部、南部、东部、沿海、西北和西南。所有这些行政区划都位于该国的南部,这是一个被几内亚的低山区森林和刚果盆地的热带雨林包围的森林地区,是潜在野生动物宿主的有利生态系统。相比之下,喀麦隆北部,一个干燥的萨赫勒和热带草原地区,似乎不太可能有利于MPXV的传播,因为在该地区没有确诊病例。由于环境干燥,该生态系统可能不适合MPXV水库。在大多数流行国家,包括塞拉利昂、尼日利亚、利比里亚、中非共和国和刚果民主共和国,mpox病例主要来自森林地区(24,25,46)。在我们的研究中,大多数MPXV确诊病例来自中部(34个[38%])和西南部(31个[25%])地区,这是该国受影响最严重的两个地区。西北地区是第三大受影响地区(18例[75%])。自2017年以来,西北和西南地区受到内乱的影响最为严重。内乱增加了该国国内流离失所者的人数,国内流离失所者经常迁移到不同的地区和邻国。此外,由于境内流离失所者在森林中的临时营地寻求庇护,这种情况大大增加了人类与野生动物的接触。由于生活在野生动物和潜在的MPXV宿主重叠的自然栖息地,西南和西北地区的种群正面临着日益增加的人畜共患MPXV感染的威胁。事实上,在非洲,内乱经常导致mpox病例增加,任何人畜共患疾病的风险都很普遍(4,49)。在几个流行国家,武装冲突或大规模人口流动背景下的mpox爆发是典型的流行病学特征,这些情况通常与低效的疾病监测和控制有关(4,49)。
基因型和系统发育分析显示,进化枝I和进化枝II同时在喀麦隆传播,并且两个进化枝之间出现明显的地理隔离。两种MPXV分支在喀麦隆的循环以前在喀麦隆的2个已发表的MPXV序列中报道过(6,29)。然而,这项研究建立在这些发现的基础上,并提供了更多的样本来进一步证实clades I和II同时在一个国家传播,这是MPXV流行病学中的一个独特特征。
在中心区域的分支II案例21V-04877中,分支的地理隔离更加明显。一项流行病学调查显示,该病例患者是一名来自西北地区的境内流离失所者,在该地区流行MPXV分支II。在喀麦隆流行的MPXV菌株之间观察到的地理隔离可归因于潜在动物宿主可能无法跨越刚果盆地热带森林覆盖的中部、东部和南部区域与几内亚低山区潮湿森林覆盖的西北和西南区域之间的天然屏障(38,40)。事实上,该国最大的河流萨纳加河和喀麦隆高原地区将这两个地理区域划分为热带潮湿森林生态区。跨萨纳加-比奥科海岸森林位于尼日利亚萨纳加河和克罗斯河之间的北部,大西洋赤道海岸森林穿过喀麦隆西南部和中非其他邻国(38,39)。或者,这两种生态环境可能拥有不同的水库。几项研究旨在识别假定的MPXV储层(31,33,47),但都没有强调特定生态系统特有的两种不同水库的潜力。此外,由于2017年以来武装冲突加剧了跨境流动,喀麦隆数十年来的人类MPXV循环可能是由于人口流动、宿主宿主宿主或两者都来自邻国的地方性宿主。新测序的MPXV菌株与来自邻近国家的对应菌株的聚类支持了这一假设,这些邻近国家与喀麦隆东部和西部没有物理屏障,但有漫长的陆地边界。
总之,这项研究提供了喀麦隆44年间mpox流行的详细信息。喀麦隆mpox的流行病学包括原发性和继发性传播。分离的进化枝I和II病毒株同时传播,表明可能存在不同的病毒库和MPXV的跨境传播。这项研究可以为喀麦隆和其他具有类似流行病学背景的非洲国家监测和控制mpox的公共卫生干预措施的设计、优化和评估提供信息。
Djuicy博士是雅温得喀麦隆巴斯德中心病毒学部门的一名研究科学家,从事人畜共患病和包括病毒性出血热在内的新出现疾病的研究。她的研究兴趣集中在为新出现和重新出现的被忽视和与贫困相关的病毒性疾病开发研究轴,包括mpox、埃博拉、拉沙热和马尔堡病毒。
感谢
我们感谢非洲疾病控制和预防中心向监测系统提供mpox诊断试剂。我们感谢同意参与本研究的患者或其法定监护人。我们感谢本研究中所有识别和收集疑似病例样本的人员。我们感谢Huguette Simo对手稿的技术审查,Landry Messanga对原始系统发育树的推断,以及Hornela Ossombo对QGIS绘图的贡献。
这项工作得到了喀麦隆巴斯德中心的支持。
参考
- 马伦尼科娃 圣人, 莫耶 辐射武器(radiation weapon的缩写)。痘病毒的分类和该属的简要特征正痘病毒属。In: Shchelkunov SN,Marennikova SS,Moyer RW,编辑。对人类致病的正痘病毒。波士顿:斯普林格;2005。第11–8页。
- 王菲 O, 屁股 民用波段, 王菲 M, 秋天 G, 奇蒂 是, 迪亚涅 abbr. 毫米(millimeter), 以及其他人 尼日利亚人猴痘病毒的基因组特征。 柳叶刀传染病. 2018;18:246. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 中泽友秀 Y, 莫尔丁 先生, 爱默生 标距长度, 雷诺兹 机关枪, 讽刺 乡邮投递路线, 高 J, 以及其他人 非洲猴痘的系统地理学调查。 病毒. 2015;7:2168–84. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 麦科勒姆 调幅;振幅调制(amplitude modulation), 达蒙 反向动力学. 人类猴痘。 临床感染疾病. 2014;58:260–7. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 里穆安 (Aw)人名, 穆伦巴卡尼 首相, 约翰斯顿 南卡罗来纳州, 劳埃德·史密斯 乔, 基萨鲁 日本海事协会, 金凯拉 元素铊的符号, 以及其他人 在刚果民主共和国天花疫苗接种运动停止30年后,人类猴痘发病率显著增加。 美国国家科学院院刊. 2010;107:16262–7. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 拉奥 阿拉斯加, 彼得森 频带宽度(Band Width), 怀特希尔 F, Razeq JH, 艾萨克斯 塞内加尔, 默赫林斯基 玛帝脂接缝(MasticJoint的缩写), 以及其他人 JYNNEOS(活的、非复制的天花和猴痘疫苗)用于正痘病毒职业暴露危险人群的暴露前疫苗接种:美国免疫实践咨询委员会的建议,2022。 Morb莫尔布凡人周代表. 2022;71:734–42. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- european theatre of operations 欧洲战区 A, 斋藤 T, 横手 H, 库拉内 I, 金谷 Y. 减毒细胞培养天花疫苗LC16m8的研究进展。 疫苗. 2015;33:6106–11. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 辛普森 K, 海曼 D, 褐色的 特许测量员, 埃德蒙兹 WJ, 埃尔斯加德 J, 很好 P, 以及其他人 人类猴痘——40年后,根除天花的意外后果。 疫苗. 2020;38:5077–81. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 贝松贝斯 C, 姆布伦加 F, 谢弗 L, 马拉卡 C, 戈诺菲奥 E, 兰迪尔 J, 以及其他人 2001-2021年中非共和国国家猴痘监测。 急诊传染病. 2022;28:2435–45. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 非洲疾病控制和预防中心。疫情简报24:非洲联盟成员国的mpox引用于2023年11月23日]. https://Africa CDC . org/disease-outbreak/outbreak-brief-24-mpox-in-Africa-union-member-States外部连接
- 埃雷兹 N, 阿奇杜特 H, 米尔罗特 E, 施瓦茨 Y, 维纳井 Y, 帕拉恩 N, 以及其他人 2018年以色列输入性猴痘的诊断。 急诊传染病. 2019;25:980–3. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 尼日利亚 仇恨失控, 李 V, 马里穆图 K, 瓦索 S, 陈 G, 林(姓氏) 实时位置(Real-TimePosition的缩写), 以及其他人 新加坡一例输入性猴痘。 柳叶刀传染病. 2019;19:1166. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 旺市 A, 阿伦斯 E, 阿斯特伯里 J, 布鲁克斯 T, 钱德 M, 弗莱格 P, 以及其他人 猴痘病毒的人际传播,英国,2018年10月。 急诊传染病. 2020;26:782–5. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 旺市 A, 阿伦斯 E, 阿斯特伯里 J, 巴拉瑟加拉姆 S, 比兹沃斯 M, 贝克 中国国家铁路(China Railway), 以及其他人 2018年9月输入英国的两例猴痘。 欧洲监察. 2018;23:1800509. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 勇 SEF, 尼日利亚 仇恨失控, 嗬 ZJM, 马克 铥, 马里穆图 K, 瓦索 S, 以及其他人 新加坡进口猴痘。 急诊传染病. 2020;26:1826–30. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 芦苇 (窑内)烘干的(Kiln-dried), 梅尔斯基 JW, 格雷姆 医学学士, 雷杰尼里 RL, 索蒂尔 玛帝脂接缝(MasticJoint的缩写), 韦格纳 市场价值(Market Value), 以及其他人 在西半球人类中发现猴痘。 英国医学杂志. 2004;350:342–50. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 疾病控制和预防中心。2022–2023年Mpox爆发全球地图[引用于2023年5月9日]. https://www . CDC . gov/pox virus/mpox/response/2022/world-map . html
- 卡勒 J, 侯赛因 A, 弗洛勒斯岛 G, 赫伊里 S, 脱rosiers D. 猴痘:传播、发病机制和表现的综合综述。 库雷乌斯. 2022;14:e26531. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 奎纳 加拿大, 摩西 C, 门罗 碱基对, 中泽友秀 Y, 多蒂 分线盒, 休斯 厘米, 以及其他人 刚果民主共和国农村社区猴痘的假定危险因素。 公共科学图书馆一号. 2017;12:e0168664. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 纳库恩 E, 兰帕特 E, 恩贾普 副检察长, 扬森斯 C, 祖尼加 I, 范赫普 M, 以及其他人中非共和国医院内人类猴痘爆发。公开论坛传染疾病。2017;4:ofx168。
- 因卡-奥贡勒耶 A, 阿鲁纳 O, 达尔哈特 M, 奥吉纳 D, 麦科勒姆 A, 苏迪 Y, 以及其他人; 疾控中心猴痘爆发小组. 2017-18年尼日利亚爆发人类猴痘:一项临床和流行病学报告。 柳叶刀传染病. 2019;19:872–9. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 利科斯 调幅;振幅调制(amplitude modulation), 萨蒙斯 救世军(Salvation Army), 奥尔森 弗吉尼亚烟草, Frace 调幅;振幅调制(amplitude modulation), 里 Y, 奥尔森-拉斯姆森 M, 以及其他人 两个分支的故事:猴痘病毒。 j基因病毒. 2005;86:2661–72. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 宽松外衣 C, 阿迪提法 I, 姆巴拉 P, 恩朱姆 R, 纳库恩 E, 宽松外衣 A, 以及其他人 急需一个非歧视性和非污名化的猴痘病毒命名法。 公共科学图书馆生物学. 2022;20:e3001769. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 伊西德罗 J, 博尔赫斯 V, 杂色的 M, 索布拉尔 D, 桑托斯 法学博士(Jurum Doctor), 努内斯 A, 以及其他人 2022年猴痘病毒多国爆发中的系统发育学特征和微进化迹象。 Nat Med. 2022;28:1569–72. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- Sadeuh-Mba 救世军(Salvation Army), 永加 机关枪, 抗剪弹性极限 M, 贝特贾特 C, 埃扬戈 S, 卡罗 V, 以及其他人 2018年在喀麦隆发现的人类病例与2017-2018年在尼日利亚爆发的猴痘病毒之间的系统发育相似性。 感染遗传进化. 2019;69:8–11. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 贝尔特 N, 德索普-德克莱 S, 贝松贝斯 C, 库拉索多 M, 恩吉利·梅扬 嗜酒者互诫协会, 塞莱孔 B, 以及其他人 2001年至2018年间中非共和国人类猴痘感染的基因组历史。 Sci代表. 2021;11:13085. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 拉多尼奇 A, 梅茨格 S, 达布罗斯基 女警, 库阿西-海曼 E, 舒纳德尔 L, 库尔特 A, 以及其他人 2012年科特迪瓦野生乌白眉猴的致命猴痘。 急诊传染病. 2014;20:1009–11. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 库拉索多 M, 贝松贝斯 C, 纳库内 E, 囟门 A, 格塞恩 A, 哈桑宁 A. 基于生态位比较识别猴痘病毒最可能的哺乳动物宿主。 病毒. 2023;15:727. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 乔科特 脉波频率(Pulse Frequency的缩写), 日本轿子 I, 泰坦耶 E, 恩多姆比 P, 皮农 D, Mbede J. 【天花还是水痘重症?喀麦隆一名儿童因猴痘病毒引起人类天花一例] [法语]. 安足球俱乐部. 1991;71:123–8.PubMed外部连接谷歌学术外部连接
- Eozenou P. 喀麦隆联合共和国一例猴痘的回顾性调查 [法语]. 公牛队。 1980;2:23–6.
- 海曼 D. 莫隆杜埃博拉-猴痘调查的初步报告(喀麦隆,1980年2月) [法语]. 公牛队。 1980;7:58–60.
- 公共卫生应急行动协调中心。喀麦隆。猴痘[法语]引用于2023年5月9日]. https://www . ccousp . cm/urgences-sanitaires/Mpox/situation-Mpox-Cameroun外部连接
- 莫卢阿 埃尔, 兰比 厘米。喀麦隆的气候、水文和水资源。比勒陀利亚:南非比勒陀利亚大学非洲环境经济学和政策中心;2006.
- 泽巴泽·托古埃 嘘, 尼亚姆西·恰乔 N, 塔尔姆 兹就, 捕鱼车 C。萨纳加河,中非喀麦隆生物物理和社会文化一体化的范例。In: Wantzen KM,编辑。河流文化:生命随着水的节奏起舞。巴黎:联合国教育、科学及文化组织;2023.
- 一个地球。非洲营养引用于2023年5月9日]. https://www.oneearth.org/realms/afrotropics外部连接
- 里 Y, 赵 H, 威尔金斯 K, 休斯 C, 达蒙 反向动力学. 猴痘病毒西非和刚果盆地株DNA的实时PCR检测。 病毒学方法. 2010;169:223–7. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 迈耶 H, Ropp 海平面(sea level), 埃斯波西托 姐姐(网络用语)ˌ法官ˌ裁判员(judges)。 A型包涵体蛋白基因可用于聚合酶链式反应分析,以区分正痘病毒。 病毒学方法. 1997;64:217–21. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 诺伦 致死剂量, 奥萨代贝 L, 卡通巴 J, 利科法塔 J, 穆卡迪 D, 门罗 B, 以及其他人 刚果民主共和国猴痘爆发期间的人与人之间的传播。 急诊传染病. 2016;22:1014–21. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 奥吉纳 D, Iroezindu M, 詹姆斯 嗨, 奥拉多昆 R, 因卡-奥贡勒耶 A, 若间 P, 以及其他人 尼日利亚人猴痘的临床过程和结果。 临床感染疾病. 2020;71:e210–4. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 拉韦 landingzone登陆区, 阿莫阿代 M, 布拉格 L, 阿德武夷 P, 香农河 F, 威尔逊 W, 以及其他人 2017年6月利比里亚Rivercess县Doedain区爆发猴痘。 流行病学公共卫生杂志。 2021;4:8. 数字对象标志符外部连接谷歌学术外部连接
- 贝松贝斯 C, 戈诺菲奥 E, 科纳姆纳 X, 塞莱孔 B, 授予 R, 格塞恩 A, 以及其他人 2018年中非共和国猴痘病毒的家庭内传播。 急诊传染病. 2019;25:1602–4. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 雷诺兹 机关枪, 多蒂 分线盒, 麦科勒姆 调幅;振幅调制(amplitude modulation), 奥尔森 弗吉尼亚烟草, 中泽友秀 Y. 猴痘在非洲重新出现:呼吁扩大同一健康的概念和实践。 专家版本抗感染治疗. 2019;17:129–39. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- Tchatchueng-Mbougua 分线盒, 梅桑加·埃森格 陆上通信线(Land Lines), Septoh Yuya 平头接合(flush joint), Kamtchogom V, 哈马杜 A, 萨德厄姆拜 救世军(Salvation Army), 以及其他人 在资源有限的环境下用数字平台改善冠状病毒肺炎19诊断测试数据的管理和安全性:喀麦隆的案例。 PLOS数字健康. 2022;1:e0000113. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
- 麦克帕克 B, 没完没了地唠叨琐事 S, 萨利 S, Wurie H, 纳马库拉 J, 森古巴 F. 受冲突影响的国家和卫生系统中的埃博拉病毒:乌干达北部和塞拉利昂的案例研究。 冲突健康. 2015;9:23. 数字对象标志符外部连接PubMed外部连接谷歌学术外部连接
图
表
原始出版日期:2024年2月7日
1目前隶属于:马里兰州农业部,美国马里兰州索尔兹伯里。
Hits: 50