
Antigen Processing and Presentation
为了能够发挥适应性免疫的关键要素(特异性、记忆性、多样性、自身/非自身识别),抗原必须经过加工处理并呈递给免疫细胞。抗原呈递由主要组织相容性复合体(MHC)I 类分子和 II 类分子介导,这些分子存在于抗原呈递细胞 (APC)以及某些其他细胞的表面。
MHC I 类和 II 类分子功能相似:它们将短肽递送至细胞表面,分别使这些肽能够被 CD8+ (细胞毒性)T 细胞和 CD4+ (辅助性)T 细胞识别。区别在于这些肽的来源不同——MHC I 类分子递送的肽来源于内源性或细胞内 ;而 MHC II 类分子递送的肽来源于外源性或 细胞外 。此外,还存在所谓的交叉呈递 ,即外源性抗原也可以通过 MHC I 类分子呈递。当内源性抗原通过自噬降解时,也可以通过 MHC II 类分子呈递。
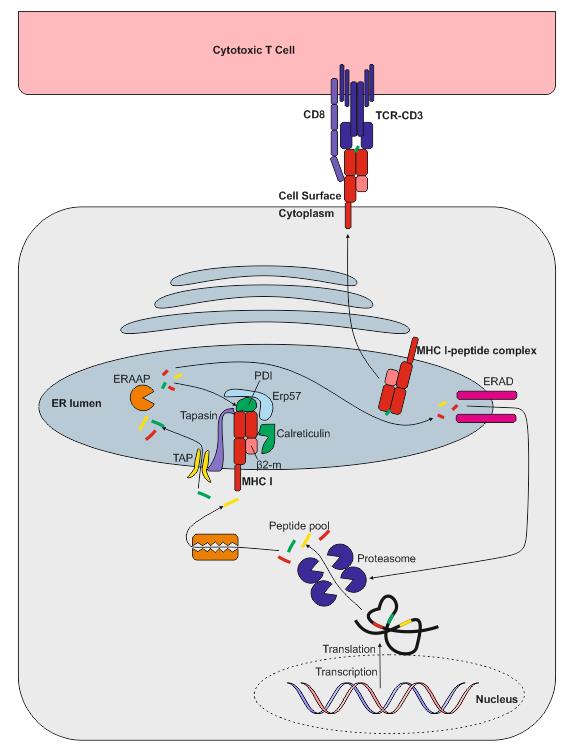
MHC I 类呈递
MHC I 类分子在所有有核细胞中均有表达。MHC I 类分子在内质网 (ER)中组装,由两种链组成:一条多态性重链和一条称为β2-微球蛋白的链。重链在与β2-微球蛋白结合之前,由分子伴侣钙网蛋白(calnexin) 稳定。在没有肽的情况下,这些分子由分子伴侣蛋白稳定,这些分子伴侣包括:钙网蛋白(calreticulin)、Erp57、蛋白二硫键异构酶(PDI)和 tapasin。TAP、tapasin、MHC I 类分子、ERp57 和钙网蛋白组成的复合物称为肽加载复合物 (PLC)。Tapasin 与转运蛋白 TAP(抗原呈递相关转运蛋白)相互作用,后者将肽从细胞质转运到内质网。在进入内质网之前,肽来源于蛋白质的降解,这些蛋白质可能来源于病毒或自身。蛋白质的降解由胞质和核蛋白酶体介导,产生的肽段通过 TAP 转运至内质网。TAP 转运 8-16 个氨基酸的肽段,这些肽段在与 MHC I 类分子结合前可能需要在内质网中进行额外的修饰。这可能是由于内质网氨肽酶(ERAAP)的存在,该酶与抗原加工相关。
值得注意的是,30%至 70%的蛋白质在合成后会立即降解(它们被称为 DRiPs——缺陷核糖体产物,是转录或翻译缺陷的结果)。这一过程使得病毒肽能够快速呈递——例如,流感病毒感染后约 1.5 小时即可被 T 细胞识别。当肽与 MHC I 类分子结合时,分子伴侣会被释放,肽-MHC I 类复合物离开内质网,呈递至细胞表面。在某些情况下,肽无法与 MHC I 类分子结合,必须返回胞质溶胶进行降解。一些 MHC I 类分子从未与肽结合,它们也会被内质网相关蛋白降解(ERAD)系统降解。
有几种不同的蛋白酶体可以生成用于 MHC I 类分子呈递的肽: 26S 蛋白酶体 ,由大多数细胞表达; 免疫蛋白酶体, 由许多免疫细胞表达;以及胸腺上皮细胞表达的胸腺特异性蛋白酶体 。
抗原呈递
在单个细胞表面,MHC I 类分子能够反映多达 10,000 种蛋白质的表达水平。细胞毒性 T 淋巴细胞和自然杀伤细胞能够解读这些信息,从而监测细胞内部的活动,并检测感染和肿瘤的发生。
细胞表面的 MHC I 类复合物会随着时间的推移而解离,其重链可被内吞。当 MHC I 类分子被内吞进入内体后,它们便进入 MHC II 类分子呈递途径。部分 MHC I 类分子可以被回收利用,并作为交叉呈递过程的一部分,呈递内体肽段。
通常情况下,抗原通过 MHC I 类分子呈递的过程基于 T 细胞受体与结合在 MHC I 类分子上的肽段之间的相互作用。此外,T 细胞表面的 CD8+ T 细胞分子也与 MHC I 类分子上的非肽段结合区域发生相互作用。因此,与 MHC I 类分子结合的肽段只能被 CD8+ T 细胞识别。这种相互作用是所谓的“三信号激活模型”的一部分,实际上是第一个信号。第二个信号是抗原呈递细胞(APC)上的 CD80/86 与 T 细胞表面的 CD28 之间的相互作用,随后是第三个信号——APC 产生细胞因子,从而完全激活 T 细胞,使其产生特异性免疫应答。
MHC I 类多态性
人类主要组织相容性复合体(MHC)I 类分子由一系列基因编码,包括 HLA-A、HLA-B 和 HLA-C(HLA 代表“人类白细胞抗原”,是大多数脊椎动物中 MHC 分子的人类对应物)。这些基因具有高度多态性,这意味着每个人都拥有其独特的 HLA 等位基因组合。这些多态性的后果是,不同个体对感染和自身免疫性疾病的易感性存在差异,这可能是由于不同个体中能够与 MHC I 类分子结合的肽段种类繁多所致。此外,MHC I 类分子的多态性使得供体和受体之间几乎不可能实现完全匹配的组织,因此也是导致移植排斥反应的原因。
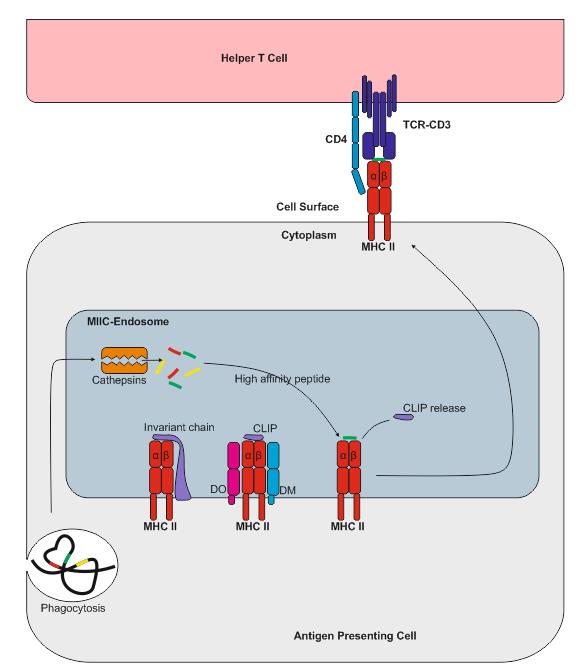
MHC II 类呈递
MHC II 类分子由抗原呈递细胞 (APC) 表达,例如树突状细胞 (DC)、 巨噬细胞和 B 细胞 (在 IFNγ 刺激下,间充质基质细胞、成纤维细胞和内皮细胞,以及上皮细胞和肠神经胶质细胞也会表达)。MHC II 类分子与内吞途径降解的蛋白质衍生的肽段结合。MHC II 类复合物由 α 链和 β 链组成,它们在内质网中组装,并由不变链 (Ii) 稳定。MHC II 类分子和 Ii 的复合物通过高尔基体转运至一个称为 MHC II 类区室 (MIIC) 的区室。由于酸性 pH 值,蛋白酶组织蛋白酶 S 和组织蛋白酶 L 被激活并消化 Ii,在 MHC II 类分子的肽结合槽中留下残留的 II 类相关 Ii 肽段 (CLIP)。随后,CLIP 被内吞途径降解的蛋白质衍生的抗原肽所取代。这一过程需要分子伴侣 HLA-DM,在 B 细胞的情况下,还需要 HLA-DO 分子。负载外源肽的 MHC II 类分子随后被转运至细胞膜,并将抗原呈递给 CD4+ T 细胞。此后,MHC II 类分子介导的抗原呈递过程基本与 MHC I 类分子介导的抗原呈递过程相同。
与 MHC I 类分子不同,MHC II 类分子不会在质膜上解离。控制 MHC II 类分子降解的机制尚未完全阐明,但 MHC II 类分子可以被泛素化,然后通过内吞途径进入细胞。
MHC II 类多态性
与 MHC I 类重链一样,人类 MHC II 类分子由三个多态性基因编码:HLA-DR、HLA-DQ 和 HLA-DP。不同的 MHC II 类等位基因可用作多种自身免疫性疾病的遗传标记,这可能与其呈递的肽类有关。
Hits: 57